Кроветворение (гемоцитопоэз) - многостадийный процесс последовательных клеточных превращений, приводящий к образованию зрелых клеток периферической сосудистой крови. В постэмбриональный период у животных развитие клеток крови осуществляется в двух специализированных интенсивно обновляющихся тканях, относящихся к разновидностям тканей внутренней среды и условно названных миелоидной и лимфоидной. В них постоянно совершается сбалансированный процесс новообразования и гибели клеточных элементов. Представлены они многочисленными гемопоэтическими клетками разного типа в комплексе с ретикулярными иди эпителиальными элементами и макрофагами.
В миелоидной ткани (греч. миелос - мозг) красного костного мозга происходит развитие стволовых кроветворных клеток и всех форменных элементов крови - эритроцитов, гранулоцитов, лимфоцитов, кровяных пластинок, моноцитов.
В лимфоидной ткани, находящейся в тимусе, селезенке, лимфатических узлах, слизистых оболочках внутренних полостных органов, образуются лимфоциты, а также клетки, являющиеся конечными стадиями дифференциации стимулированных T- и B-лимфоцитов.
С помощью клональных, иммунологических, электронно-микроскопических, генетических и радиобиологических методов за последние 25 лет получены важные данные, характеризующие кинетику клеточных популяций в процессе кроветворения. Отражением этого явилось построение новых схем кроветворения, в которых уточнены ранние стадии гемоцитопоэза, когда разделение клеток по морфологическим признакам еще невозможно. В настоящее время наиболее признанной является схема кроветворения, предложенная И. Л. Чертковым и А. И. Воробьевым (1981), в соответствии с которой весь гемоцитопоэз разделен на шесть этапов и соответственно выделено шесть классов кроветворных клеток (цв. табл. II).
Исходя из представления, сформулированного более 60 лет назад А. А. Максимовым, о происхождении клеток крови из единого источника признано, что родоначальным элементом всех клеток крови является полипотентная стволовая клетка (колониеобразующая единица в селезенке - КОЕс), способная к разнообразным превращениям и обладающая свойствомсамоподдержания (пролиферации без видимой дифференциации) своего численного состава в течение всей жизни организма. Популяция стволовых клеток в схеме кроветворения считается клетками первого класса. Во взрослом организме наибольшее количество стволовых клеток находится в красном костном мозге (на 100000 клеток костного мозга приходится около 50 стволовых), из которого они мигрируют в тимус, селезенку, а у птиц в фабрициеву сумку.
Пролиферируют и развиваются стволовые клетки в том или ином направлении под влиянием близкодействующих индукторов микроокружения, образуемых клетками стромы - различными клетками ретикулярной (в красном костном мозге, селезенке) или ретикуло-эпителиальной основы (в тимусе). Несмотря на то что стволовая клетка кроветворения способна проделывать около 100 митозов, в нормальных физиологических условиях основная масса стволовых клеток митотически инертна. Усиление их митотической активности и восстановление характерного для кроветворной системы данного организма количества стволовых клеток происходят при воздействиях, резко снижающих общую массу гемопоэтических элементов (например, после кровопотерь или воздействия лучистой энергии). Светомикроскопическое и электронно-микроскопическое исследование наиболее очищенной фракции стволовых клеток показало, что они имеют по своей морфологии сходство с малыми лимфоцитами.
Ближайшей ступенью превращения стволовой клетки в процессе кроветворения является второй класс - частично детерминированных клеток-предшественников двух разновидностей - миелопоэза и лпмфопоэза. Это популяция полустволовых клеток с более ограниченными способностями к самоподдержанию. На агаровой культуре эти клетки образуют колонии, поэтому они получили название "колониеобразующие единицы" - КОЕ. Подтверждено существование клетки-предшественницы гранулоцитарного, эритроцитарного, моноцитарного и мегакариоцитарного рядов гемопоэза (КОЕ - ГЭММ). Интенсивность их размножения и превращения в следующий, третий класс - "унипотентные клетки-предшественницы", обладающие еще меньшими способностями к самоподдержанию, регулируется действием специфических биологически активных веществ - поэтинов.
В настоящее время в третий класс поэтинчувствительных клеток отнесены клетки, способные к дифференцировке в направлении как двух ростков - клетка-предшественница грануло- и моноцитопоэза (КОЕ - ГМ), клетка гранулоцито- и эритроцитопоэза (КОЕ - ГЭ), клетка мегакариоцито- и эритроцптопоэза (КОЕ - МГЦЭ), так и клетки, дифференцирующиеся лишь в одном направлении, - клетка-предшественница гранулоцитов (КОЕ - Г), клетка-предшественница моноцитопоэза (KOE-M), клетка-предшественница эозинофилов (КОЕ - Эо), клетка-предшественница базофилов (КОЕ - Б), клетка-предшественница мегакариоцитов (КОЕ - МГЦ). Что касается лимфопоэза, то еще не полученоподтверждения существования общей (для T- и В-лимфоцитов) клетки-предшественницы, и она в схеме остается гипотетичной. Однако на основании обнаружения соответствующих клеточных антигенных маркеров выявлены клетки-предшественницы отдельно для T- и В-лимфоцитов.
Перечисленные выше классы стволовых, полустволовых и унипотентных предшественников имеют лимфоцитоподобный вид и морфологическими методами не распознаются. Если за счет стволовых клеток происходит качественная регуляция кроветворения, то есть снабжение кроветворной системы всеми видами предшественников, то на стадии поэтинчувствительных и следующих за ней морфологически распознаваемых стадиях большинство клеток находится в состоянии пролиферации. Именно в этом отделе реализуется основная количественная регуляция кроветворения, то есть обеспечение необходимого количества клеток нужного типа в ответ на конкретные потребности организма.
Далее следует четвертый класс клеток типа "бластов" (эритробласты, миелобласты, лимфобласты и т. д.). Все они имеют более крупные размеры (20 мкм и более), ядро с нуклеолами и нежносетчатым хроматином, неширокий ободок беззернистой, слабобазофильпой цитоплазмы. Несмотря на то что каждый "бласт" развивается в направлении лишь одного определенного типа клеток, морфологически все они трудно различимы.
Пятый и шестой классы морфологически распознаваемых клеток - это соответственно класс созревающих (миелоцит, нормоцит и др.) и класс зрелых клеток (эритроциты, гранулоциты и др.). На уровне последних двух классов выявлено принципиальное различие в поведении клеток миелоидного и лимфоидного рядов. Если в последних стадиях миелоидного кроветворения развитие идет вплоть до гибели клеток, то в лимфоидном ряду возможно превращение морфологически зрелых лимфоцитов в бластные формы. Однако это происходит под влиянием специфических индукторов - антигенов (антигензависимая бласттрансформация). Таким образом, в основном подтверждается выдвинутое А. А. Максимовым (1927) представление о том, что малый лимфоцит крови не является конечной стадией дифференциации клеток лимфоидного ряда, а сохраняет способность трансформироваться в клетки, способные к митотическому делению.
Развитие эритроцитов (эритроцитопоэз) в красном костном мозге протекает по схеме: стволовая клетка (CK) - полустволовые клетки (КОЕ - ГЭММ, КОЕ - ГЭ, КОЕ - МГЦЭ) - унипотентные предшественники эритропоэза (БОЕ - Э, КОЕ - Э) - эритробласт - пронормоцит - нормоцит базофильный - нормоцит поли-хроматофильный - нормоцит оксифильный - ретикулоцит - эритроцит. До стадии эритробласта клеткам несвойственны характерные отличительные морфологические признаки, как полагают, они имеют лимфоцитоподобный вид. О свойствах этих клеток судят на основании данных, получаемых главным образом методом клонирования в полутвердых средах, содержащих агар, метилцеллюлозу и др. Показано, что в обычных условиях эритроцитопоэза непосредственный предшественник - эритропоэтинчувствительная унипотентная клетка (КОЕ - Э) образуется из клеток, формирующих большие колонии - бурсты, состоящие из нескольких тысяч эритроидных клеток, так называемые бурстообразующие единицы (БОЕ - Э). В условиях повышенной потребности в эритроцитах эритроцитопоэз может миновать стадии БОЕ - Э и КОЕ - Э (на схеме обозначено пунктирными линиями).
Конечный период эритроцитопоэза (начиная с эритробластов) сопровождается образованием морфологически распознаваемых клеток. При этом происходят характерные морфологические изменения: уменьшаются размеры всей клетки, отмечают ее уплотнение, затем исчезает ядро, изменяется окраска цитоплазмы. Эритробласты - клетки размером от 15 до 25 мкм. Ядро, занимающее ее большую часть, содержит мелко распыленный хроматин и 1 - 3 ядрышка. Образующиеся из эритробластов пронормоциты имеют меньшие размеры (12 - 18 мкм) и более грубую структуру хроматина ядра (рис. 101). Пронормоциты - интенсивно делящиеся клетки. В результате митотического деления образуются клетки размером 10 - 12 мкм, с плотным ядром и интенсивно
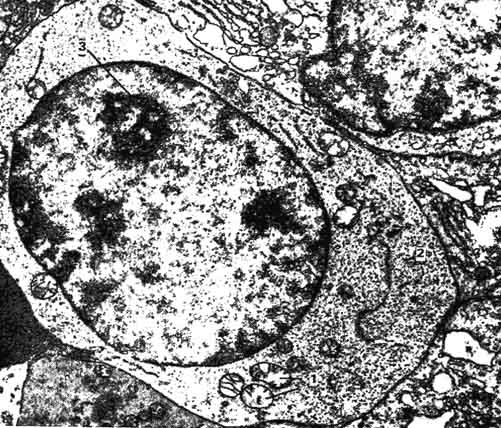
Рис. 101. Электронная микрограмма пронормоцита (по Агееву):
1 - митохондрии; 2 - рибосомы; 3 - ядрышко.
базофильной цитоплазмой, в которой становится заметной узкая более светлая перинуклеарная зона - базофильные нормоциты.
Базофилия цитоплазмы обусловлена наличием в ней большого количества РНК, свободных рибосом и полисом, с которыми связан синтез белкового компонента гемоглобина. Последний накапливается сначала в перинуклеарной зоне. Железо, входящее в состав гемоглобина, базофильные нормоциты получают от макрофагов, фагоцитирующих гибнущие эритроциты. В результате деления базофильных нормоцитов появляются еще более мелкие клетки, цитоплазма которых из-за накопленного гемоглобина утрачивает выраженную ранее базофилию и окрашивается как основными, так и кислыми красителями - полихроматофильные нормоциты. Ядра их, как правило, без ядрышек, а вследствие радиального расположения грубых глыбок гетерохроматина имеют вид колеса со спицами. Проделав завершающее деление, полихроматофильные нормоциты превращаются в клетки, цитоплазма которых вследствие обилия в ней гемоглобина проявляет выраженную оксифилию - оксифильные нормоциты. Ядро их постепенно уменьшается, пикнотизируется (уплотняется) и отсоединяется. Отделившиеся ядра нормоцитов фагоцитируются макрофагами костного мозга. Образуются молодые эритроциты - ретикулоциты, поступающие в кровяное русло. В них еще некоторое время сохраняются РНК-содержащие структуры в виде сеточки. В процессе развития морфологически распознаваемые клетки эритроцитарного ряда осуществляют 5 - 6 митозов.
Установлено, что даже в нормальных условиях кроветворения часть эритробластов (до 10%) не завершает свой цикл развития до эритроцитов и с помощью макрофагов костного мозга разрушается.
Этот процесс, названный неэффективным эритропоэзом, является одним из физиологически обусловленных механизмов регуляции в системе эритропоэза. Наиболее сильным регулятором эритропоэза является количество кислорода, доставляемого к тканям и органам. Недостаточное снабжение кислородом стимулирует усиленную выработку эритропоэтина, посредством которого регулируется интенсивность пролиферации костномозговых предшественников (преимущественно на уровне БОЕ - Э и КОЕ - Э) эритропоэза. Эритропоэтиы - гормон гликопротеидной природы. Считают, что он синтезируется в почках.
Развитие гранулоцитов (гранулоцитопоэз). При развитии гранулоцитов из стволовых клеток красного костного мозга вначале также образуются морфологически нераспознаваемые полустволовые (КОЕ - ТЭММ; КОЕ - ГМ; КОЕ - ГЭ) и унипотентные предшественники (КОЕ - Б; КОЕ - Эо; КОЕ - Гн), которые через стадии распознаваемых клеточных форм (миелобласт, промиелоцит, миелоцит, метамиелоцит, палочкоядерный гранулоцит) превращаются в зрелые сегментоядерные гранулоциты трех разновидностей - нейтрофилы, эозинофилы и базофилы.
Миелобласт - первая морфологически идентифицируемая клетка гранулоцитарного ряда - имеет крупное центрально расположенное ядро, в котором на фоне диффузно-мелкозернистого хроматина видно несколько ядрышек. Цитоплазма слабобазофильная, в ней можно обнаружить небольшое количество азурофильных гранул. Электронно-микроскопически в ней выявляются митохондрии, полирибосомы, элементы гранулярной цитоплазматической сети. Разделившись, миелобласт превращается в промиелоцит - крупную клетку (20 - 25 мкм), в которой округлое ядро часто расположено эксцентрично и содержит 1 - 2 ядрышка. В элементах пластинчатого комплекса Гольджи промиелоцитов происходит основной процесс формирования азурофильных гранул. Они в значительном количестве содержатся в цитоплазме и дают положительную реакцию на пероксидазу. После деления из промиелоцита образуются клетки еще меньших размеров и с более плотным ядром, имеющим чаще всего овальную форму, - миелоциты. В цитоплазме миелоцитов наряду с первичной (азурофильной) зернистостью образуются и содержатся вторичные (специфические) гранулы, в соответствии с особенностями которых удается отчетливо различать нейтрофильные, эозинофильные и базофильные миелоциты.
В развитии гранулоцитов миелоциты являются завершающими клетками, способными к митотическому делению, после которого они созревают, последовательно превращаясь в метамиелоциты, палочкоядерные и сегментоядерные клетки. Эти этапы созревания характеризуются некоторыми общими морфологическими изменениями: уменьшением клетки в размерах, изменением формы ядра от овальной (миелоциты) через бобовидную (метамиелоциты) и палочковидную к расчлененной на дольки (сегментоядерные) и окончательным оформлением в цитоплазме соответствующей специфической зернистости. Сегментоядерные гарнулоциты поступают из костного мозга в кровяное русло, циркулируют в нем 8 - 12 ч, после чего проникают в ткани, где выполняют специфические функции и погибают. Показано, что для эозинофилов тканевая фаза их жизни продолжается около 10 сут, базофилы же погибают очень быстро.
Развитие моноцитов (моноцитопоэз). Клетки моноцитарпого ряда образуются в костном мозге из стволовых клеток через стадии полустволовых клеток (КОЕ - ГЭММ и КОЕ - ГМ), из которых возникают унипотентный предшественник (КОЕ - M) и затем монобласты. Немногочисленные монобласты трудно отличимы от других бластных форм в красном костном мозге. После того как в их цитоплазме сформируются компоненты комплекса Гольджи и образуются азурофильные гранулы, клетки превращаются в промоноциты и моноциты. Последние выходят в кровоток, а затем, проникая в ткани, дают начало незрелым, а позднее зрелым макрофагам.
Последовательное развитие клеток линии моноцитов - макрофагов из монобластов представлено на схеме.
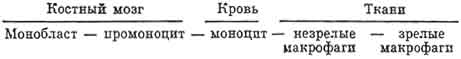
Развитие лимфоцитов. Лимфоцитопоэз - один из наиболее сложных процессов дифференцировки стволовых кроветворных клеток. Важная особенность этого процесса состоит в том, что развивается сходная морфологически, но разнородная в функциональном отношении клеточная популяция. С участием различных органов поэтапно осуществляется формирование двух тесно связанных при функционировании линий клеток - T- и В-лимфоцитов. В красном костном мозге образуются родоначальные лимфоидные клетки, общие как для T-, так и для В-лимфоцитов. В центральных лимфоидных органах (тимусе, фабрициевой сумке) лимфоцитопоэз зависит от наличия жизнеспособных костномозговых предшественников. В периферических лимфоидных органах (лимфатические узлы, селезенка, лимфоидные образования слизистых оболочек) лимфоцитопоэз является антигензависимым процессом.
Развитие кровяных пластинок (тромбоцитопоэз). Образование кровяных пластинок происходит в красном костном мозге и связано с развитием в нем особых гигантских клеток - мегакариоцитов. Мегакариоцитопоэз состоит из следующих стадий: стволовая клетка (CK) - полустволовые клетки (КОЕ - ГЭММ и КОЕ - МГЦЭ) - унипотентные предшественники (КОЕ - МГЦ) - мегакариобласт - промегакариоцит - мегакариоцит. По мере созревания, в результате своеобразной многократной эндометотпческой репродукции, формируются крупные клетки (40 - 50 мкм), содержащие в многолопастном ядре до 32 - 64 хромосомных наборов. В развивающихся мегакариоцитах, в цитоплазме образуются система микрофиламентов и микротрубочек, а также специфические гранулы. На заключительных этапах с участием формирующейся системы из гладких мембран происходит фрагментация цитоплазмы мегакариоцитов на обособленные участки - кровяные пластинки, которые через стенки синусоидов красного костного мозга попадают в кровоток. После отсоединения пластинок вокруг оставшегося ядра мегакариоцитов возникает новая цитоплазма. Полагают, что в каждом мегакариоците красного костного мозга совершается циклический процесс развития нескольких поколений кровяных пластинок.