Родоначальниками половых клеток являются первичные половые клетки - гаметобласты (гомобласты). Выявляются они микроморфологическими и цитохимическими методами в начале эмбриогенеза в стенке желточного мешка (плодовая оболочка зародыша) вблизи кровеносных сосудов. Гонобласты - крупные клетки неправильно округлой формы с большим светлым ядром, цитоплазма богата гликогеном и характеризуется высокой активностью щелочной фосфатазы. Гонобласты интенсивно делятся митозом и с током крови или по ходу кровеносных сосудов мигрируют в зачатки половых желез (гонады), где окружаются поддерживающими (фолликулярными) клетками. Последние выполняют трофическую функцию. Затем, в связи с развитием пола животного, половые клетки приобретают свойства, характерные для спермиев и яйцеклеток.
Развитие спермиев (сперматогенез) (рис. 38 - 1) протекает в семенниках (половой железе самца) половозрелого животного. В сперматогенезе различают четыре периода: размножение, рост, созревание и формирование.
Период размножения. На этой стадии клетки называются сперматогониями. Они имеют небольшие размеры, диплоидное (двойное) число хромосом, мелкие округлые ядра, богатые хроматином. Клетки интенсивно делятся митозом. Некоторые из
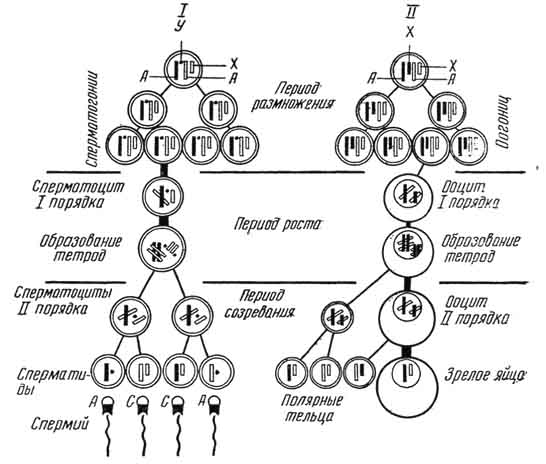
Рис. 38. Схема сперматогенеза (I) и овогенеза (II) - отцовские хромосомы черные, материнские - светлые:
А - аутосомы; Z, Y - половые хромосомы. В период размножения показаны три поколения сперматогониев и огониев, делящиеся митотически и остающиеся диплоидными.
них перестают делиться, начинают увеличиваться в размере и переходят во второй период своего развития - период роста. Другая часть спермиев продолжает делиться. Они являются стволовыми клетками и пополняют запас сперматогоний.
Период роста. В этот период клетки называются первичными сперматоцитами. У них сохраняется диплоидное числа хромосом. В процессе роста увеличивается размер клетки и происходят сложные изменения в перераспределении наследственного материала в ядре, в связи с чем различают четыре стадии: лептотенную, зиготенную, пахитенную, диплотенную.
Для лептотенной (leptos - тонкий, taenia - лента) стадии характерны хорошо выраженное ядрышко, ядерная тека и спирализация хромосом, котоые имеют вид тонких нитей. Гомологичные хромосомы (отцовские и материнские) друг с другом не конъюгируют. При зиготенной (zygoo - соединять), или синаптической (synapsis - соединение), стадии гомологичные хромосомы сближаются. Присоединившись друг к другу (конъюгировав) и образовав пары - диады, обмениваются генами - кроссинговер (crossing - скрещивание, over - через). При пахитенной (pachys - толстый) стадии пары гомологичных хромосом перекручиваются друг с другом, сильно спирализуются, поэтому укорачиваются и утолщаются. Диплотенная (diploos - двойной) стадия - процесс формирования тетрад. В каждой паре возникает продольная щель, и в двух конъюгировавших хромосомах образуются четыре хроматиды, которые начинают постепенно разъединяться, сохраняя на некоторое время связь друг с другом в центре своими концами. Затем эти связи теряются.
Период созревания. Это процесс развития сперматид с половинным числом хромосом, что достигается путем двух, следующих друг за другом делений созревания. После первого деления первичного сперматоцита образуются два вторичных сперматоцита, каждый из которых получает только половину от количества хромосом первичного сперматоцита. Вскоре за первым делением созревания без интерфазы (следовательно, без синтеза ДНК, характерного для периода S, в ходе которого увеличивается число хромосом) следует второе деление созревания. Из каждого вторичного сперматоцита образуется по две сперматиды. При этом пары хроматид, соединенные между собой в центре центромерой, разъединяются и распределяются по разным сперматидам, становясь хромосомами.
Следовательно, в процессе созревания из каждого первичного сперматоцита возникает четыре сперматиды с одинарным числом хромосом. Половые X- и У-хромосомы, характерные для спермиев, расходятся по разным сперматидам, в связи с чем Х-хромосомами снабжаются две сперматиды, другие две - У-хромосомами. Этот процесс обусловливает гетерогаметность самцов млекопитающих.
Способ деления, при котором дочерние клетки получают вдвое меньше число хромосом, носит название мейоз (meiosis - уменьшение), или редукционное (reducere - уменьшение) деление.
Сперматиды имеют округлую форму, небольшие размеры, относительно крупное ядро, бедное хроматином. Митохондрии, комплекс Гольджи, центросома хорошо развиты и расположены вблизи ядра. Другие органеллы, а также включения почти отсутствуют. Сперматиды не способны делиться.
Период формирования (рис. 39). Завершающий период сперматогенеза, в ходе которого округлая клетка - сперматида - приобретает морфологические свойства, характерные для спермия. В ходе формирования светлое округлое ядро сперматиды постепенно уплотняется, становится овальным и перемещается к плазмолемме. Комплекс Гольджи передвигается к ядру и продуцирует уплотеннную гранулу - акробласт. Последний увеличивается в размере и в виде чехлика охватывает ядро сперматиды - будущую головку спермия.
В средней зоне акробласта из мельчайших зерен формируется акросома. Это уплотненное тельце, богатое ферментом гиалуронидазой. К противоположному от ядра полюсу перемещается центросома, в которой различают проксимальную и дистальную
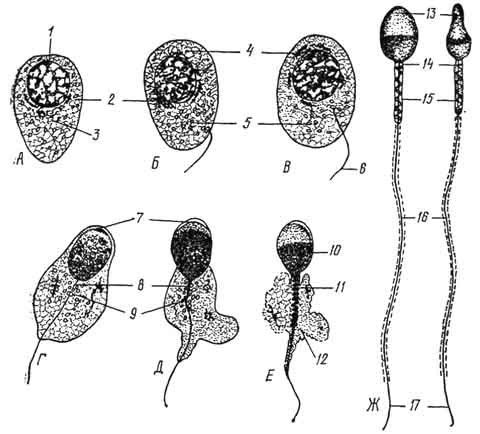
Рис. 39. Период формирования (схема). Дифференцировка сперматиды и образование спермия (А - Ж).
1 - ядро сперматиды; 2 - комплекс Гольджи; 3 - центриоли; 4 - зачаток акросомы; 5 - митохондрии; 6 - жгутик; 7 - акросомный чехлик; 8 - проксимальная центриоль; 9 - дистальная центриоль; 10 - ядерное вещество, сконцентрированное в головке спермия; 11 - митохондриальная спираль; 12 - остатки цитоплазмы; 13 - головка; 14 - шейка; 15 - средняя часть; 15 - хвостик; 17 - концевой участок хвостика.
центриоли. Они образуют шейку спермия. Дистальная центриоль делится на две части - переднюю и заднюю. От передней части дистальной центриоли отрастает осевая нить хвостика, состоящая из микротрубочек. Задняя часть дистальной центриоли приобретает вид колечка. Она, сползая по осевой нити хвостика и увлекая за собой цитоплазму сперматиды, содержащую митохондрии и гликоген, располагается на границе между начальным и главным отделами хвостика спермия. Смещаясь по хвостику, цитоплазма тонким слоем одевает главную его часть. Клетка продолжает удлиняться и приобретает форму спермия.
Развитие яйцеклеток, или овогенез (см. рис. 38 - II), - сложный и очень длительный процесс. Он начинается в период эмбриогенеза и завершается в органах половой системы половозрелой самки. Слагается овогенез из трех периодов: размножения, роста, созревания.
Период размножения протекает в период внутриутробного развития и завершается в течение первых месяцев после рождения. Клетки называются овогониями (оогониями). Они незначительных размеров, бедны цитоплазмой, их ядра имеют диплоидное число хромосом. Оогонии интенсивно митотически делятся. Некоторые из них перестают делиться, покрываются клетками фолликулярного эпителия, которые снабжают яйцеклетку питательным материалом.
В период роста клетки называются первичными ооцитами. Они увеличиваются в размере. Изменения в ядрах аналогичны первичным сперматоцитам; они проходят также лептотенную, зиготенную, пахитенную и диплотенную стадии. Затем в ооците начинается интенсивный синтез и накопление желтка, необходимого для течения первых стадий развития зародыша, в связи с чем хромосомы вновь деспирализуются. Этот процесс протекает в две фазы: стадия малого роста (превителлогенез) и стадия большого роста (вителлогенез). Морфологические особенности превителлогенеза - локализация органелл вокруг ядра, активизация функции белоксинтезирующей системы цитоплазмы: развиваются рибосомы, гранулярная эндоплазматическая сеть, поэтому цитоплазма окрашивается основными красителями, то есть она базофильна. Хорошо развиты комплекс Гольджи и митохондрии. Ядро округлое, содержит мало хроматина, хорошо развито ядрышко. Вторичная оболочка ооцита состоит из одного слоя фолликулярных клеток. Превителлогенез обычно длится до наступления половой зрелости самки.
Вителлогенез - стадия интенсивного накопления желтка в цитоплазме первичного ооцита. В данном процессе принимает участие весь организм. Фолликулярные клетки в ооцит транспортируют желток. Они интенсивно делятся митозом, поэтому вторичная оболочка становится многослойной.
Органеллы из перинуклеарной зоны перемещаются на периферию: формируется кортикальный слой, который играет важную роль в начальных стадиях эмбриогенеза.
Период созревания состоит из двух быстро следующих друг за другом делений созревания, в ходе которых диплоидная клетка становится гаплоидной. Этот процесс обычно протекает в яйцеводе после выхода первичного ооцита из яичника.
Первое деление созревания завершается образованием двух неравноценных структур - вторичного ооцита и первого направительного, или редукционного, тельца. Каждая из них по сравнению с первичным ооцитом содержит половину количества ядерного вещества. За первым делением очень скоро наступает второе, в ходе которого также образуются только одна зрелая яйцеклетка и второе неправильное тельце. Первое направительное тельце к этому моменту делится еще один раз и образуются два направительных тельца.
Следовательно, из одного первичного ооцита в процессе созревания возникают только одна зрелая яйцеклетка и три направительных тельца.
Таким образом, овогенез отличается от сперматогенеза рядом особенностей: начинается в период внутриутробного развития самки, имеет длительный период роста, в процессе которого накапливается питательный материал (желток); протекает в яичнике эмбриона и половозрелого животного, а затем в яйцеводе; из одного первичного ооцита образуется только одна зрелая яйцеклетка; все яйцеклетки генетически однородны, так как имеют Х-хромосому; женская половая клетка не способна к самостоятельному передвижению.