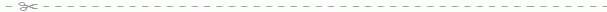Половые клетки. Оплодотворение. Дробление. Спермии птиц так же, как и у других представителей позвоночных, имеют жгутиковую форму и состоят из головки, шейки, хвостового отдела.
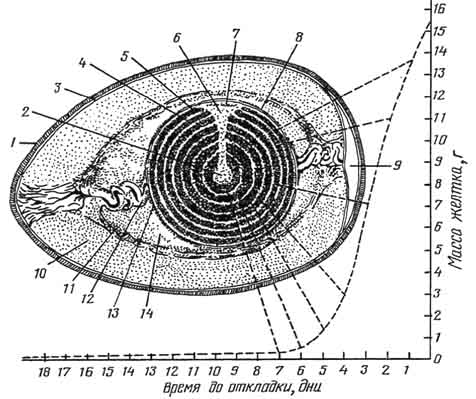
Рис. 55. Схема строения куриного яйца:
1 - скорлупа; 2 - латебра; 3 - подскорлуповая оболочка; 4 - белый желток; 5 - желтый желток; 6 - ядро Пандера (белый желток, лежащий под латеброй); 7 - бластодерма; 8 - желточная оболочка; 9 - воздушная камера; 10 - яичный белок (наружный слой альбумина); 11 - яичный белок (волокнистый слой); 12 - халаза; 13 - халазообразный слой; 14 - яичный белок (внутренний слой альбумина). Кривая справа отражает скорость роста яйца на протяжении 18 дней, предшествовавших его откладке. Штриховые линии, ведущие от разных слоев желтка к кривой роста, показывают, в какое именно время формировались эти слои.
У разных видов сельскохозяйственных птиц они отличаются размером, формой головки и хвостика. Например, верхняя часть головки спермия петуха лопастной формы, а у гусака она вытянута и заострена на конце.
Продолжительность жизни спермиев в половых путях самки более 30 дней. В течение данного периода без повторного спаривания несушки откладывают оплодотворенные яйца. В первые две недели после спаривания число оплодотворенных яиц наибольшее.
По количеству и локализации желтка яйцеклетки птиц относят к полилецитальным, телолецитальным. В яйце птиц содержатся: желток, то есть яйцеклетка, белок, подскорлуповая оболочка и скорлупа (рис. 55). Все компоненты яйца, кроме желтка, относятся к третичным оболочкам. Их материал продуцируется железами яйцевода. При продвижении яйцеклетки по яйцеводу она покрывается третичными оболочками.
Яйцеклетка, как и другие клетки, имеет ядро и цитоплазму. Ядро, расположенное в анимальном полюсе клетки, окруженонезначительным слоем цитоплазмы, содержащей органеллы. Вся остальная часть яйцеклетки заполнена желтком - клеточным включением. В состав желтка входят протеиды и липиды. В результате концентрации из растворенного состояния они осаждаются и приобретают вид шариков. В начальной стадии желточные шарики богаты фосфолипидами, а затем насыщенными жирными кислотами. У кур в желточных шариках много свободного холестерина.
Желток яйца неоднороден, он бывает светлый и темный. В центре яйцеклетки находится светлый желток. Эта зона называется латеброй и по форме напоминает колбу. В остальной части яйцеклетки темный и светлый желток расположен слоями. Обычно светлый желток откладывается в ночное время, а темный - в другое время суток.
Периферический слой цитоплазмы яйцеклетки называется кортикальным, а оболочка яйцеклетки, или плазмолемма, - первичной оболочкой. В белке яйцеклетка подвешена на халазах, направленных от желтка к тупому и острому концу яйца. Халазы - это пучки тонких скрученных волоконец, состоящих из плотного белково-углеводного комплекса.
Халазы удерживают желток в центре яйца таким образом, что зародыш всегда оказывается сверху.
Снаружи от желтка располагается белок. По месту его локализации различают наружный и внутренний слои, каждый из которых состоит из жидкого и плотного белка. В яичный белок входят вода (87%), протеины, гликопротеиды, свободные углеводы, незначительное количество липидов, золы и других веществ. Для развивающегося зародыша белок - это основной источник питательных веществ и воды среднего периода эмбриогенеза. Белок является поставщиком минеральных веществ, протеинов. Он содержит бактерицидные вещества, убивающие микроорганизмы.
Белок покрыт подскорлуповой оболочкой, в которой различают два слоя: наружный и внутренний. Наружный слой толщиной около 56 мкм состоит из волокон диаметром 2 - 10 мкм. Этот слой плотно соединен со скорлупой. Толщина внутреннего слоя равна 16 мкм, а диаметр ее волокон 2 - 3 мкм. По химическому составу волокна сходны с кератиноподобным (роговым) веществом. Внутренний слой подскорлуповой оболочки прилегает к белку.
В зоне тупого конца яйца слои подскорлуповой оболочки расходятся и образуется воздушная камера, содержащая воздух.
Подскорлуповая оболочка очень плотна, эластична и проницаема для газов, воды, растворимых соединений. Свойства оболочки в значительной степени зависят от ее влажности. Влажная подскорлуповая оболочка набухает, в ней увеличиваются поры, роль которых выполняют пространства между волокнами.
Содержимое яйца заключено в скорлупу. Последняя защищает яйцо от повреждений, ее материал используется зародышем в качестве минеральных веществ при построении скелета. Скорлупаимеет поры, через которые происходят газообмен и испарение влаги при развитии зародыша.
В состав скорлупы входят органические и минеральные вещества. Из органических веществ построены плотно сплетенные волокна и гранулы белка. Снаружи скорлупа покрыта тонкой надскорлуповой пленкой - кутикулой. Построенная из муцина, она препятствует проникновению через поры скорлупы микроорганизмов, спор грибов и проницаема для газов.
После выхода из яичника яйцеклетка попадает в яйцевод, где она оплодотворяется. При спаривании самца с самкой p яйцевод попадает одновременно несколько миллионов спермиев. Особенностью оплодотворения яйцеклеток сельскохозяйственных птиц является полиспермия - в яйцеклетку может проникать до 300 спермиев, с ядром женской половой клетки соединяется только один спермий. После оплодотворения, продвигаясь по яйцеводу и покрываясь третичными оболочками (белком, подскорлуповой оболочкой, скорлупой), яйцеклетка интенсивно делится митозом. Следовательно, снести яйцо - это зародыш на ранней стадии эмбриогенеза.
Дробление у птиц меробластическое (частичное) или дискоидальное. В нем принимает участие только анимальный полюс зиготы, где расположены ядро и цитоплазма, лишенная желтка. Вегетативный полюс не участвует в дроблении, так как он загружен желтком, тормозящим дробление.
Первые две борозды дробления имеют меридиональное направление и расположены перпендикулярно друг другу. Меридиональные борозды сменяются широтными (рис. 56). Образовавшиеся при дроблении клетки (бластомеры) лежат на желтке в виде одного слоя - диска, поэтому дробление получило название дискоидальнего, а образовавшаяся при этом бластула - дискобластулы. Ее крыша и краевая зона состоят из бластомеров диска, а дно - из неразделенного на клетки желтка.
Бластоцель имеет вид узкой щели и сдвинут к анимальному полюсу бластулы (рис. 57).
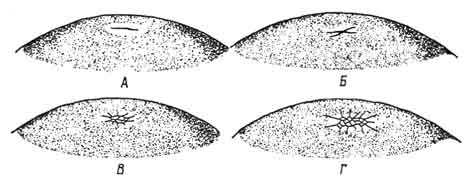
Рис. 56. Дробление куриного яйца:
А - 2 бластомера; Б - 4 бластомера; В - 8 бластомеров; Г - около 16 бластомеров.
Рис. 57. Зародышевый диск куриного яйца на поздней стадии дробления (А) и на стадии, близкой к гаструляции (Б).
1 - бластомеры; 2 - эктодерма; 3 - бластоцель; 4 - желток.
После оплодотворения яйцо в яйцеводе может находиться в течение 4 - 27 ч, поэтому и дробление, и гаструляция протекают в этом органе.
В снесенном яйце эмбриогенез приостанавливается и вновь возобновляется с момента начала инкубации или насиживания.
Гаструляция. Закладка осевых органов. Развитие зародышевых листков. Через 12 ч инкубации в центре зародышевого диска образуется зародышевый щиток. Из этой части зародышевого диска развивается тело зародыша. Остальная часть диска называется незародышевой, так как она используется при построении плодных оболочек (временных, или провизорных, органов), обеспечивающих нормальное развитие цыпленка. Зародышевый щиток окружен светлым полем. Его клетки приподняты над желтком. Под ними располагается подзародышевая полость, образовавшаяся вследствие использования зародышем желтка. Светлое поле окружено темным полем. Его клетки интенсивно делятся и разрастаются по поверхности желтка. Эту зону называют краем обрастания.
Гаструляция у птиц, как и у других позвоночных, протекает в две фазы. Первая фаза: путем расслоения, или деляминации, однослойного диска образуются два зародышевых листка - эктодерма и энтодерма (рис. 58). Вторая фаза - формирование хордомезодермального зачатка: клетки краевой зоны диско-бластулы интенсивно делятся и мигрируют в направлении заднего края зародышевого щитка двумя потоками, где они встречаются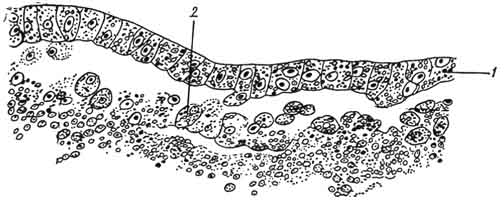
Рис. 58. Гаструляция у птиц на ранней стадии. Деляминация и эмиграция:
1 - эктодерма; 2 - энтодерма.
и начинают продвигаться вперед по средней линии зародышевого щитка, формируя утолщенный клеточный валик - первичную полоску. Она направлена от заднего края зародышевого щитка вперед.
В средней зоне первичной полоски образуется углубление - первичная бороздка (рис. 59), на переднем конце которой возникает утолщение. Это первичный, или гензеновский, узелок. Он имеет углубление, носящее название первичной ямки (гастрального мешочка).
Первичная полоска идентична бластопору гаструлы ланцетника и амфибий. Первичная ямка гензеновского узелка - это гомолог дорсальной губы бластопора, остальные зоны первичной полоски гомологичны боковым и вентральной губам.
В зоне первичной ямки зародышевый материал инвагинирует и в виде клеточного тяжа перемещается между эктодермой и энтодермой к переднему концу зародыша. Этот тяж называется головным, или хордальным, отростком, из него развивается хорда (рис. 60).
Через первичную бороздку в направлении переднего конца зародыша мигрируют два крылообразных зачатка. Они растут с боков от хорды между эктодермой и энтодермой.

Рис. 59. Гаструляция у птиц:
А - миграция клеток в зародышевом щитке; Б - образование хордомезодермального зачатка; 1 - эктодерма; 2 - материал будущей нервной пластинки; 3 - материал хордальной пластинки; 4 - первичный (головной) узелок; 5 - первичная ямка; 6 - первичная полоска; 7 - первичная бороздка; 8 - хорда; 9 - мезодерма. Сплошные стрелки обозначают направление перемещения материала в составе наружного, а пунктирные - в составе среднего зародышевого листка (по Кнорре).
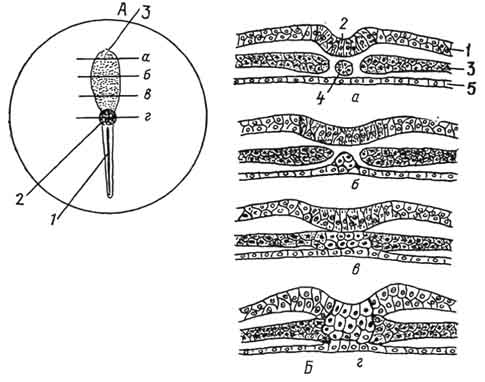
Рис. 60. Изменения в зародышевом щитке при гаструляции (А):
1 - первичная полоска, образованная мезодермальным материалом; 2 - первичный (головной) узелок, образованный зачатковым материалом хорды; 3 - зачаток нервной пластинки; Б - четыре поперечных разреза гаструлы, сделанные на уровнях, которые указаны на рисунке А (а - через передний конец тела, б, в - через средний отдел тела, г - первичный узелок); 1 - кожная эктодерма; 2 - нервная пластинка; 3 - мезодерма; 4 - хорда; 5 - кишечная энтодерма.
Дифференцировка этих зачатков заканчивается образованием среднего зародышевого листка - мезодермы.
Таким образом, и у птиц гаструляция завершается закладкой зародышевых листков. В гаструляции птиц отмечают дальнейшее усложнение в развитии позвоночных: зачаток хорды и мезодермы полностью утратили связь с первичной кишкой. Последняя очень рано отделяется от других эмбриональных зачатков и входит в состав энтодермы зародышевого щитка.
Дальнейшее развитие зародышевых листков у птиц протекает, как и у других позвоночных (см. рис. 53). Из эктодермы выделяется нервная пластинка, а после соединения ее краев формируется нервная трубка. Эктодерма наползает снаружи на нервную трубку, в связи с чем последняя оказывается погруженной под эктодерму. Нервная трубка - это источник развития всей нервной системы, а эктодерма - зачаток поверхностного слоя кожного покрова (эпидермиса).
Мезодерма подразделяется на сомиты (сегментированную мезодерму) , сегментные ножки, несегментированную мезодерму (спланхнотом). В состав сомитов входят дермотом, миотом, склеротом. Из дермотома развиваются глубокие слои кожного покрова, из миотома - мышечная ткань скелета, из склеротома - скелет. Сегментные ножки - источник развития мочевыделительной системы.
В спланхнотоме различают париетальный (наружный) и
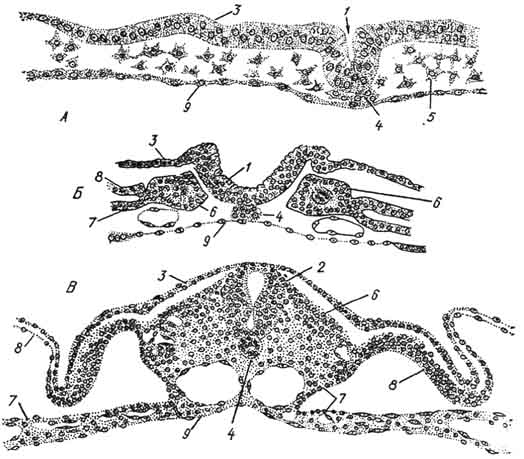
Рис. 61. Образование осевых зачатков у зародыша курицы:
А, Б, В - три последовательные стадии; 1 - нервный желобок; 2 - нервная трубка; 3 - эктодерма; 4 - хорда; 5 - мезодерма; 6 - сомиты; 7 - висцеральный листок спланхнотома; 8 - париетальный листок спланхнотома; 9 - кишечная энтодерма.
висцеральный (внутренний) листки, между которыми находится целом - вторичная полость тела. Из спланхпотома развивается эпителиальная ткань, покрывающая серозные оболочки внутренних органов, грудную и брюшную полости (рис. 61).
Образование внезародышевых органов. Развитие тела зародыша сопровождается образованием временных, или внезародышевых (провизорных), органов, способствующих созданию необходимых условий для нормального развития зародыша. К провизорным органам у птиц относят: желточный мешок, амнион, серозу, аллантоис. Они построены также из зародышевых листков - эктодермы, энтодермы, листков несегментированной мезодермы. Источником развития этих листков является внезародышевая зона зародышевого диска (цв. табл. I).
Темпы развития временных органов часто могут опережать развитие зародыша, у птиц по сравнению с другими животными этот процесс протекает более интенсивно.
Образование провизорных органов начинается с разрастания по поверхности желтка периферической зоны внезародышевого материала бластодиска. Эта зона называется зоной обрастания, после завершения гаструляции она состоит из внезародышевых эктодермы, энтодермы и листков несегментированной мезодермы.
В дальнейшем, при развитии плодных оболочек, эктодерма всегда связана с париетальным листком, а энтодерма - с висцеральным листком мезодермы.
Ранее других плодных оболочек образуется желточный мешок, формирующийся в процессе обрастания желтка зародышевыми листками. Он выполняет трофическую функцию и построен из внезародышевых энтодермы и висцерального листка мезодермы. Энтодерма желточного мешка участвует в ферментативном расщеплении желтка и всасывании питательных веществ в кровеносные сосуды, расположенные в его мезодермальном слое. Так как яйцеклетка птиц содержит большое количество желтка, эта плодная оболочка развита очень хорошо. Желточный мешок отшнуровывается от тела зародыша туловищной складкой. Ее образование обусловлено интенсивным разрастанием клеток всех зародышевых листков, лежащих на границе зародышевого и незародышевого материала. В туловищной складке участвуют все зародышевые листки этой зоны. Острие складки направлено под туловище зародыша, в связи с чем зародыш приподнимается над желтком, а его туловищная энтодерма, ранее распластанная на желтке, образует кишечную трубку. Последняя сообщается с желтком посредством узкого желточного протока.
Туловищная складка первоначально развивается в головной части зародыша, а затем распространяется в каудальном направлении.
Амнион (amnion - чаша), или водная оболочка. Сероза. Одновременно с туловищной складкой, отграничивающей зародышевую от незародышевой части, образуется амниотическая складка. Ее гребень направлен над телом зародыша. Возникая сначала над головой зародыша, она постепенно захватывает туловище. Головная часть амниотической складки образуется очень быстро, поэтому ее стенка состоит только из эктодермы. После срастания краев головной части амниотической складки возникает проамнион. Туловищная зона амниотической складки отстает в развитии. В ее состав входит не только эктодерма, но и париетальный листок мезодермы.
Над туловищем зародыша края амниотической складки срастаются: эктодерма с эктодермой, париетальный листок с париетальным листком мезодермы. После срастания краев амниотической складки тело зародыша оказывается окруженным двумя плодными оболочками - амнионом и серозой.
В формировании амниона принимают участие эктодерма и париетальный листок мезодермы внутренней половины амниотической складки, второй - эктодерма и париетальный листок наружной половины амниотической складки. Края хвостовой части амниотической складки не срастаются, здесь образуется канал, связывающий полость амниона с остальной частью яйца. Этотканал получил название серо-амниотического канала. Эктодерма амниона продуцирует амниотическую жидкость, которая заполняет амниотическую полость, создавая для развивающегося зародыша самую благоприятную водную среду. Амнион выполняет защитную функцию, он сглаживает удары и создает для эмбриона возможность некоторой подвижности.
Сероза - сама поверхностная оболочка зародыша. Прилегая к скорлупе, она покрывает снаружи амнион с расположенным в нем зародышем и желточный мешок с желтком.
Основными функциями серозы является участие в газообмене и переносе ионов кальция из скорлупы в тело зародыша. Электронно-микроскопическое строение клеток эктодермы серозы сходно с клетками, продуцирующими хлориды: плазмолемма снабжена многочисленными микроворсинками, в цитоплазме очень много митохондрий. Эти клетки синтезируют соляную кислоту, действие которой направлено, очевидно, на растворение солей кальция скорлупы, которые затем переводятся в организм плода.
Аллантоис (allantois - колбасообразный) образуется после завершения формирования кишечника на вентральной поверхности задней кишки. Он, как и кишечник, состоит из энтодермы и висцерального листка мезодермы. Аллантоис постепенно заполняет все щели между амнионом, желточным мешком и серозой, срастаясь с мезодермальным листком последней. В этом участке формируется густая сеть кровеносных сосудов, что способствует улучшению снабжения организма кислородом.
Таким образом, прилегая к скорлупе, аллантоис участвует в газообмене. Он выполняет функцию выделительного органа, так как в нем скапливаются продукты обмена веществ.
Все провизорные органы в конце эмбриогенеза редуцируются. Из приведенного материала следует, что развитие птиц также протекает стадийно. Нарушение типичного течения одной из стадий влечет за собой изменения всего хода развития зародыша. Знание стадийности развития позволяет выделить наиболее важные периоды в формировании зародыша. Понимание их крайне необходимо в связи с задачами практического птицеводства.
Однако в настоящее время периодизация в развитии птиц еще недостаточно разработана. Наиболее детально изучено развитие куриного зародыша.
H. П. Третьяков и M. Д. Попов, основываясь на характере питания зародыша, разработали следующую классификацию стадий развития цыпленка.
1-я стадия - латебральное питание - протекает первые 30 - 36 ч. Материалом для питания служит желток латебры, содержащий белки, соли, воду. Источником энергии является гликоген, поэтому зародыш почти не нуждается в кислороде. В организме отсутствует кровообращение.
2-я стадия - желточное питание и наличие желточного круга кровообращения - длится с 30 - 36 ч до 7 - 8 дней инкубации. К 30-му часу инкубации закладываются сердце и желточныймешок, в последнем развиваются кровеносные сосуды. Запас гликогена почти полностью исчезает. В организм поступает кислород, что облегчает использование белков и жиров. Формируются и другие плодные оболочки, а также органы тела: развивается нервная система, начинается биение сердца и сокращение мускулатуры тела. Закладка печени обусловливает начало синтеза мочевины, поэтому продукты распада белков становятся менее вредными для зародыша.
3-я стадия - дыхание кислородом воздуха и питание белком яйца - продолжается с 7 - 8-го дня по 18 - 19-й день. К концу ее зародыш становится сформированным плодом. Для этой стадии характерны интенсивное развитие и функционирование аллантоиса, в стенке которого формируется густая сеть кровеносных сосудов. Прилегая к серозе и вместе с ней к подскорлуповой оболочке яйца, аллантоис обеспечивает снабжение кислородом развивающиеся ткани. Благодаря растворению скорлупы возрастает интенсивность минерального обмена, усиливается усвоение жиров.
4-я стадия - потребление кислорода воздуха из воздушной камеры яйца. Цыпленок переходит на легочное дыхание, начинает функционировать малый круг кровообращения, появляется артериальная кровь, обогащающая ткани кислородом. Продолжается стадия с 18 - 19-го дня до наклева. К этому моменту аллантоис подвергается обратному развитию, и цыпленок начинает испытывать недостаток в кислороде. Он проклевывает внутренний листок подскорлуповой оболочки и начинает дышать воздухом, содержавшимся в воздушной камере яйца.
5-я стадия - вылупление - длится с 20-х по 21-е сутки. Цыпленок питается остатками желтка, которые вместе с желточным мешком втягиваются в полость кишки. Другие зародышевые оболочки также прекращают функционировать и отмирают. Цыпленок проклевывает скорлупу и покидает яйцо.
Существуют и другие классификации. Согласно разработанной Г. А. Шмидтом и M. H. Рогозиной периодизации, основанной на эмбриогенезе куриного зародыша, различают четыре периода развития: зародышевый, предплодный, плодный, а также период вылупления.
Зародышевый период длится первые восемь дней, из которых начальные стадии развития протекают во время продвижения зародыша по яйцеводу. Зародышевый период характеризуется интенсивно протекающими процессами формообразования. Источником питания зародыша является желток и частично смешанная с ним жидкая фракция белка. После образования желточного мешка транспортировка питательных веществ осуществляется с помощью его кровеносных сосудов. Дыхание зародыша сначала протекает осмотическим путем, а затем обеспечивается сосудистой системой желточного мешка. В конце зародышевого периода в этом процессе принимают участие кровеносные сосуды аллантоиса.
Предплодный период начинается с конца 8-го дня и завершается к 13 суткам инкубации. Все провизорные органы достигают функциональной зрелости. Наряду с основным внекишечным способом питания зародыша, который осуществляется посредством желточного мешка, имеет место усвоение материала внутрикишечным способом: цыпленок заглатывает амниотическую жидкость из полости амниона.
Дыхание предплода осуществляется исключительно через сосудистую систему аллантоиса.
Плодный период охватывает срок с 13 - 14 суток до начала 20-х суток инкубации. Характерным для этого периода является интенсивный рост органов тела плода. Основной способ питания зародыша - это внутрикишечный, внекишечный способ становится дополнительным. Белок, переместившийся в полость амниона и смешавшийся с амниотической жидкостью, является источником питания. Сохраняется аллантоидное дыхание. Кровеносные сосуды аллантоиса находятся вблизи подскорлуповой оболочки. Это способствует усилению газообмена, что необходимо в связи с интенсивным развитием органов тела цыпленка.
Период вылупления - на 20 - 21-е сутки инкубации. Он характеризуется прекращением функций временных органов, кишечным пищеварением, дыханием с участием легких.
Таким образом, данные о приспособительных реакциях организма и о закономерностях формообразования, свойственные каждому периоду эмбриогенеза, являются научным обоснованием при разработке и внедрении мероприятий, направленных на дальнейшее совершенствование технологии искусственного выведения сельскохозяйственных птиц.