Скелетная мышечная ткань - это сократительная ткань туловища, головы, конечностей, глотки, гортани, верхней половины пищевода, языка, жевательных мышц. Данную ткань относят к произвольной мускулатуре, так как ее сокращение контролируется волей животного.
Скелетная мышечная ткань развивается из миотомов сегментированного отдела мезодермы, а исчерченная мышечная ткань внутренних органов - из спланхнотома.
На ранней стадии развития миотомы состоят из плотно расположенных мышечных клеток - миобластов. Это первая стадия гистогенеза - миобластическая. Цитоплазма миобластов имеет тонковолокнистое строение, свидетельствующее о развитии сократительных белков. Уже на этой стадии миобласты способны к сокращению. Они интенсивно делятся и перемещаются клеточными потоками в участки расположения будущих мышц (рис. 138). Вскоре в цитоплазме миобластов можно различить единичные сократительные нити - миофибриллы, построенные из сократительных белков. Ядра миобластов относительно крупные, овальные, с малым количеством гетерохроматина и
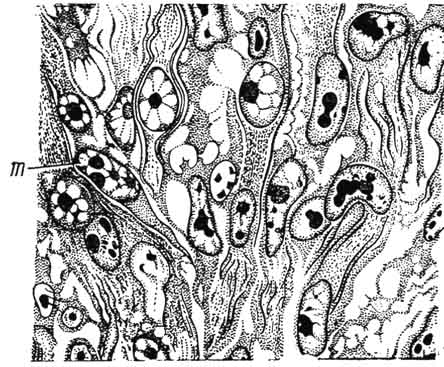
Рис. 138. Дифференцировка миобластов (т) в потоке клеток, выселяющихся из миотома.
хорошо выраженными ядрышками. Они делятся интенсивнее клеток, поэтому вскоре миобласты становятся многоядерными. Увеличиваясь в длину, они приобретают форму волокон - симпластов.
В центре симпласта в ряд располагаются многочисленные ядра, на периферии интенсивно дифференцируются миофибриллы. Миосимпласты, очевидно, могут образовываться и путем слияния миобластов. Это вторая стадия гистогенеза. Она называется стадией мышечных трубочек. Мышечные трубочки, расщепляясь вдоль, формируют мышечные волокна. В последних число миофибрилл резко увеличивается, многочисленные ядра перемещаются на периферию и располагаются под плазмолеммой. Волокно приобретает исчерченность. Это третья стадия гистогенеза - стадия мышечных волокон. К мышечным волокнам подрастает соединительная ткань с кровеносными сосудами, нервами, дифференцируются нервные окончания. Соединительная ткань участвует в образовании наружной оболочки мышечного волокна и связывает между собой мышечные волокна (рис. 139).
Сведения о гистогенезе помогут понять строение скелетной мышечной ткани и разобраться в тех сложных изменениях, которые протекают в ней при физической нагрузке, тренировке, в условиях физиологической регенерации и патологии.
Процесс регенерации, протекающий в скелетной мышечной ткани, сходен с гистогенезом; в нем выявлены те же
миобластическая стадия, стадия мышечных трубок и стадия мышечных волокон.
Как следует из гистогенеза, у дифференцированной скелетной мышечной ткани нет клеточного строения. Ее структурной и функциональной единицей является мышечное волокно (рис. 140) в виде длинных цитоплазматических тяжей с закругленными концами, которые могут переходить в сухожилия. Длина волокон 10 - 100 мкм. Состоит мышечное волокно из саркоплазмы (цитоплазмы) и многочисленных ядер, расположенных на периферии. Само волокно покрыто сарколеммой (оболочкой). Структурные компоненты саркоплазмы - сократительный аппарат, органеллы, включения, гиалоплазма. Понять механизм сокращения скелетной мышечной ткани можно лишь после знакомства с тончайшей структурной организацией всех его компонентов.
Сократительным аппаратом скелетного мышечного волокна являются продольно ориентированные миофибриллы. Построенные из сократительных белков, они занимают большую часть волокна, оттесняя ядра на периферию. Диаметр
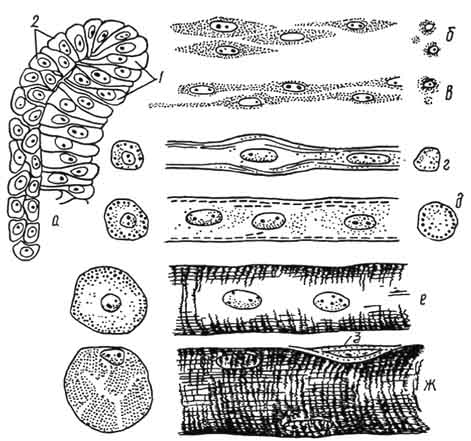
Рис. 139. Основные этапы эмбриогенеза скелетно-мышечной ткани:
а - клетки сомита (1 - миотом, 2 - дермотом); б - миобласты; в - миосимпласты; г - промиотуба; д - мышечная трубочка; е - незрелое мышечное волокно; ж - зрелое мышечное волокно; 3 - клетка соединительной ткани. Стадии б - ж показаны на продольном и поперечном разрезах.
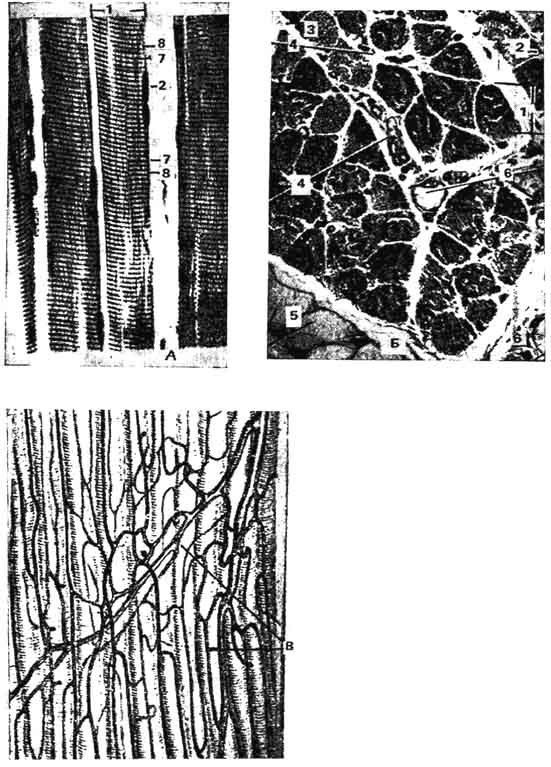
Рис. 140. Исчерченная скелетная мышечная ткань:
А - продольное сечение; Б - попереченое сечение; 1 - мышечное волокно; 2 - ядро мышечного волокна; 3 - миофибриллы; 4 - соединительная ткань перимизия; 5 - жировые клетки; 6 - кровеносный сосуд; 7 - анизотропный диск; 8 - изотропный диск; В - кровеносные сосуды мышечных волокон.
миофибрилл около 1 - 2 мкм. Миофибриллы состоят из чередующихся темных и светлых полос (дисков). Все светлые и все темные диски миофибрилл в одном мышечном волокне удерживаются на одном уровне, в связи с чем волокно приобретает поперечную исчерченность. Продольная ориентация миофибрилл

Рис. 141. Строение миофибрилл исчерченной скелетной мышечной ткани:
А - диск (анизотропный); I - диск (изотропный); Z-линия (телофрагма); М -линия (мезофрагма) (по Хаксли). Электронная микрофотография.
может создавать продольную исчерченность мышечного волокна.
В поляризованном свете темные полосы (диски) обнаруживают двойное лучепреломление - анизотропию, поэтому их называют анизотропными, или полосами А (дисками А). Светлые полосы изотропны, их называют изотропными, или полосами I (дисками I). В середине каждого I диска проходит темная зона - линия Z (телофрагма). В середине А диска проходит светлая зона - линия H с темной линией посередине - линией M (мезофрагма) (рис. 141), Диски и линии были открыты очень давно с помощью оптического микроскопа. Они хорошо видны на изолированных миофибриллах, которые можно получить, расщепив мышечное волокно.
Структурной единицей миофибриллы является саркомер. В миофибрилле они расположены, следуя друг за другом. Саркомер - это участок миофибржллы, состоящий из линия Z (для двух соседних саркомеров), половины диска I, диска А с линией Н, половины следующего диска I1 линии Z (для двух соседних саркомеров). Эти компоненты миофибрилл связывали с сокращениями, однако их участие в этом процессе оставалось неясным. Электронно-микроскопические, гистохимические, биохимические исследования многое внесли в расшифровку функциональной морфологии саркомера. Было установлено, что диск А состоит из более толстых (диаметром 10 нм, длиной 1,5 мкм) миофиламентов, диск I - из более тонких (диаметром 5 нм, длиной 1 мкм) миофиламентов. Материалом для построения толстых миофиламентов служит белок миозин, а тонких - актин, тропомиозин В, тропин.
Актиновые и миозиновые миофиламенты контактируют не конец в конец, а перемещаются по отношению друг к другу и в диске А образуют зону перекрытия. Участок А диска, состоящий только из миозиновых миофиламентов, называется H линией и по сравнению с зоной перекрытия более светлый. Линия M - это место соединения толстых миозиновых миофиламентов в анизотропном диске.
Линия Z состоит из Z-филаментов. В них выявлены белки тропомиозин-В, а-актин. Z-филаменты формируют решетку, к
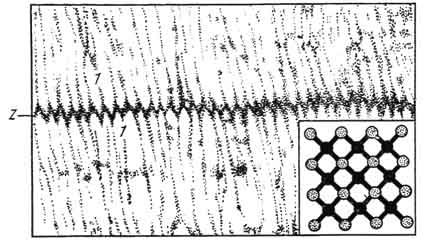
Pиc. 142. Линия Z:
1 - прикрепление к ней тонких миофиламентов. Вставка внизу поясняет прикрепление тонких миофиламентов к Z. Электронная микрофотография.
которой с обеих сторон прикреплены тонкие актиновые филаменты полосок I двух соседних саркомеров. Z линия проходит через всю толщину саркомера, а зона прикрепления тонких миофиламентов имеет зигзагообразный контур (рис. 142).
Таким образом, линии Z и M являются опорным аппаратом саркомера.
В строении сократительного аппарата при сокращении мышечного волокна наблюдаются следующие изменения: уменьшается длина саркомеров, так как тонкие (актиновые) миофиламенты полоски I при скольжении между толстыми (миозиновыми) филаментами полоски А сдвигаются к линии M диска А. Это приводит к увеличению зоны перекрытия, формированию боковых мостиков между актиновыми и миозиновыми миофиламентами (рис. 143), сокращению линий H, сближению линий Z (рис. 144).
В гиалоплазме мышечного волокна хорошо развиты митохондрии - органеллы клеточного дыхания. Они скапливаются между миофибриллами, вокруг многочисленных ядер, вблизи сарколеммы, то есть в тех зонах, которые характеризуются значительной потребляемостью АТФ. Этим объясняется высокая метаболическая активность скелетного мышечного волокна.
Интенсивное развитие в мышечном волокне имеет незернистая эндоплазматическая сеть (саркоплазматический ретикулум). Ее мембранные элементы расположены вдоль саркомеров и в виде терминальных цистерн окружают линии Z (рис. 145). Саркоплазматической сети присуща специфическая функция накопления ионов кальция, необходимых при сокращении и расслаблении мышечного волокна.
Остальные органеллы (зернистая эндоплазматическая сеть, комплекс Гольджи и др.) развиты слабее и локализуются около ядер.
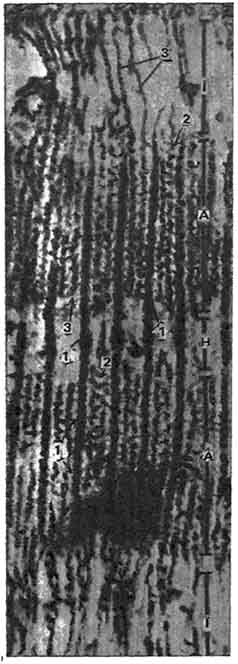
Рис. 143. Участок саркомера исчерченной мышечной ткани:
1 - толстые миофиламенты; 2 - поперечные мостики; 3 - тонкие миофиламенты. А - 1/2 диска A; I - 1/2 диска I; H - зона, состоящая только из толстых миофиламентов (по Хаксли).
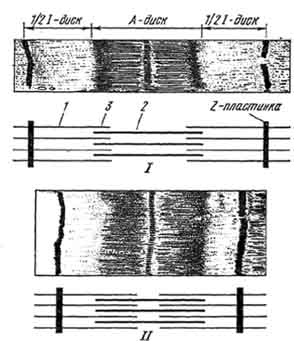
Рис. 144. Саркомер поперечнополосатого мышечного волокна в расслабленном (I) и сокращенном состоянии (II):
1 - тонкие нити; 2 - толстые нити; 3 - зона перекрытия.
Между миофибриллами находится значительное количество гранул гликогена (трофического) включения - материала для синтеза АТФ.
В цитоплазме мышечного волокна содержатся дыхательные ферменты, белок, миоглобулин - аналог гемоглобина эритроцитов; последний также способен связывать ж отдавать кислород.
В мышечном волокне ядра расположены на периферии вблизи сарколеммы. Они имеют овальную форму и варьируют по количеству от десяти до нескольких сотен. Гетерохроматин в виде крупных глыбок находится в относительно светлой нуклеоплазме. Ядра могут располагаться в виде цепочки, следуя друг за
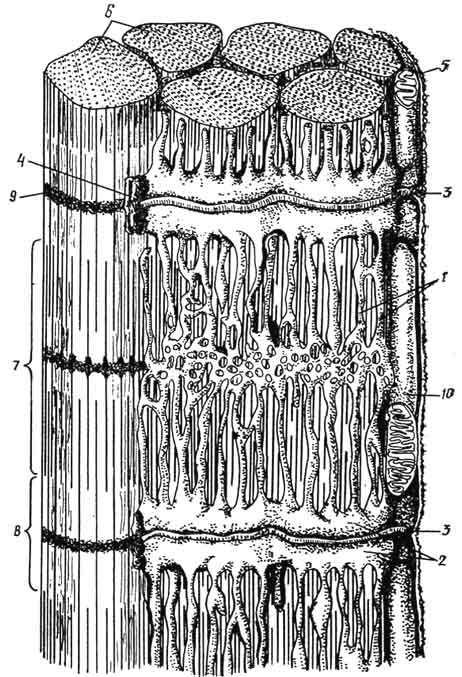
Рис. 145. Схема участка исчерченного мышечного волокна:
1 - саркоплазматическая сеть; 2 - терминальные цистерны саркоплазматической сети; 3 - Т-трубка; 4 - триада; 5 - сарколемма; 6 - миофибриллы; 7 - диск А; 8 - диск I; 9 - линия; Z; 10 - митохондрии.
другом. Это результат амитотического деления - показатель реактивного состояния мышечного волокна.
Снаружи мышечное волокно покрыто оболочкой - сарколеммой, состоящей из внутреннего и наружного слоев. Внутренний слой - это плазмолемма, которая аналогична оболочке других тканевых клеток. Наружный - соединительнотканный слой
194
<состоит из базальной мембраны и прилегающих к ней волокнистых структур. Плазмолемма образует систему узких канальцев, проникающих внутрь мышечного волокна. Это система поперечных трубочек (Т-система). У млекопитающих системы Т-трубок расположены снаружи саркомеров на границе А и I дисков. У других классов животных она проникает в волокно на уровне линии Z. Золы контакта системы поперечных трубок, саркоплазматической сети и терминальных цистерн называются триадами. Они играют главную роль в продвижении волн деполяризации и аккумуляции ионов кальция. Триады видны только в электронный микроскоп.
Плазмолемма мышечного волокна, как и нервных волокон, электрически поляризована. В расслабленном мышечном волокне на ее внутренней стороне поддерживается отрицательный, на наружной стороне - положительный потенциал.
При мышечном сокращении волна деполяризации по нервному волокну через нервное окончание перемещается на плазмолемму мышечного волокна, вызывая ее местную деполяризацию. Через систему T-трубок, связанную с плазмолеммой, и триаду волна деполяризации оказывает влияние на проницаемость мембран саркоплазматической сети, побуждая высвобождение в саркоплазму аккумулированных в ней ионов кальция. В присутствии последних активируется расщепление АТФ, что необходимо для образования актомиозинового комплекса и скольжения актиновых миофиламентов по отношению миозиновых миофиламентов. Это вызывает укорочение каждого саркомера, а следовательно, миофибрилл и мышечных волокон в целом.
Важное место в этом процессе занимают молекулы толстых мпофиламентов - миозина. Эти молекулы состоят из головки я длинного хвостика. Они при гидролизе АТФ, чему способствует АТФ-азная активность головок молекул миозина, вступают в связь с определенными участками молекул тонких миофиламентов - актином (см. рис. 143). Тонкие нити сдвигаются к центру саркомера, линии Z сближаются, увеличиваются зоны перекрытия, сокращаются линии H анизотропных дисков миофибрилл (см. рис. 144). Затем с участием АТФ актомиозиновые связи разрушаются, а миозиновые головки присоединяются к соседним участкам актиновых нитей, что способствует дальнейшему продвижению миофиламентов по отношению друг к другу.
Если концентрация ионов кальция в саркоплазме снижается и они перекачиваются в саркоплазматическую сеть, то сокращение мышечного волокна прекращается. Для этого процесса также необходима АТФ. Следовательно, и при сокращении, и при расслаблении мышечного волокна расходуется АТФ, источником которой являются глюкоза, гликоген, жирные кислоты.
Сарколемма на концах скелетных мышечных волокон формирует пальцеобразные выросты. Между ними находятся коллагеновые волокна соединительной ткани фасций и сухожилий, прикрепляющие волокна к скелету.
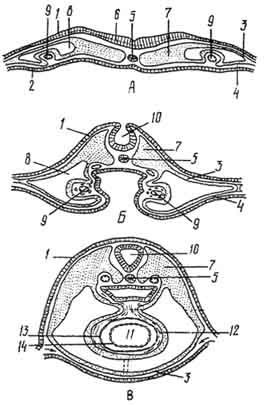
Рис. 146. Развитие сердца:
А - В - поперечные разрезы зародышей на трех послестадиях формирования трубчатой закладки сердца; А - две парные закладки сердца; Б - их сближение; В - их слияние в одну непарную закладку; 1 - эктодерма; 2 - энтодерма; 3 - париетальный листок мезодермы; 4 - висцеральный листок; 5 - хорда; 6 - нервная пластинка; 7 - сомит; 8 - вторичная полость тела; 9 - эндотелиальная закладка сердца (парная); 10 - нервная трубка; 11 - полость сердца; 12 - эпикард; 13 - миокард; 14 - эндокард.
Соединительнотканные волокна, расположенные снаружи базальной мембраны мышечного волокна, образуют эндомизий, который богат кровеносными сосудами и нервами. Эндомизий соединяется с перимизием - оболочкой, покрывающей группу мышечных волокон. Перимизий нескольких мышечных пучков соединяется с эпимизием - самой наружной соединительнотканной оболочкой, объединяющей несколько таких пучков в мышцу - орган, характеризующийся специфическим строением и функцией.
Сердечная мышечная ткань. Этот вид мышечной ткани образует среднюю оболочку сердца, по характеру сокращения относится к непроизвольной, так как не контролируется волей животного. Развивается она из участка висцерального листка мезодермы - миоэпикардиальной пластинки. Свое название эмбриональный зачаток получил в связи с тем, что из него развивается и другая оболочка сердца - эпикард (рис. 146).
Сердечная мышечная ткань состоит из мышечных клеток - кардиомиоцитов (сердечных миоцитов). Миоциты, соединяясь друг с другом своими концами по длинной оси клеток, формируют структуру, сходную с мышечным волокном (рис. 147). Границы между соседними миоцитами - это вставочные диски - аналоги линий Z, которые имеют прямые или ступенчатые контуры. Вставочные диски обеспечивают механическую прочность мышечного пласта и электрическую связь между кардиомиоцитами.
Различия в строении и функции миоцитов дали основание классифицировать сердечную мышечную ткань на две разновидности: рабочую и проводящую. Первая составляет большую часть сердечной мышцы.
Кардиомиоциты на своей поверхности несут отростки или анастомозы, так как с их помощью клетки соединяются друг с другом. Сердечные миоциты - это одноядерные и реже
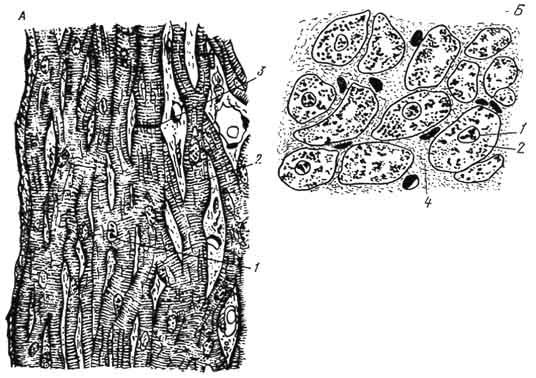
Рис. 147.
Сердечная мышечная ткань (А - продольное и Б - поперечное сечение):
1 - ядро; 2 - цитоплазма клетки; 3 - вставочные полоски; 4 - рыхлая соединительная ткань.
двуядерные клетки. Их светлые ядра овальной формы расположены в центре клетки. Цитоплазма (саркоплазма) состоит из сократительных нитей - миофибрилл, органелл, включений и гиалоплазмы. Органеллы клеток локализуются у полюсов ядра. Хорошо развиты митохондрии, хуже комплекс Гольджи, саркоплазматическая сеть. Включения представлены многочисленными гранулами гликогена и пигмента липофусцина. Количества последнего увеличиваются пропорционально возрасту.
Сократительный аппарат миоцитов так же, как в скелетной мышечной ткани, состоит из миофибрилл, которые занимают периферическую часть клетки. Их диаметр варьирует от 1 до 3 мкм. По своему строению миофибриллы сходны с таковыми скелетной мышечной ткани. Они также построены из анизотропных (полосы А) и изотропных (полосы I) дисков. Этим обусловлена их поперечная исчерченность (рис. 148).
Элементы саркоплазматической сети окружают миофибриллы. Характерное свойство сердечных миоцитов - отсутствие терминальных цистерн, а поэтому и триад.
Плазмолемма кардиомиоцитов на уровень Z линий инвагинирует в глубь цитоплазмы, образуя поперечные трубочки (Т-систему). От скелетной мышечной ткани они отличаются большим диаметром и наличием базальной мембраны, которая, как и сарколемму, покрывает их снаружи. Волны деполяризации, идущие с плазмолеммы, а также по Т-системе внутрь сердечных миоцитов, вызывают скольжение актиновых миофиламентов по отношению миозиновых, обусловливая сокращение, как и в скелетной мышечной ткани.
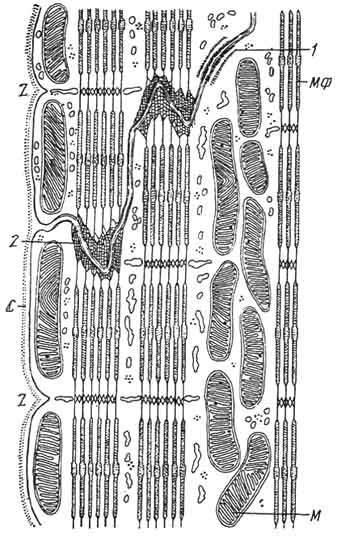
Рис. 148. Схема строения сердечной мышцы в области ступенчатой вставочной полоски:
С - сарколемма; M - митохондрии; МФ - миофиламенты; 1 - зона уплотнения на клеточной оболочке; 2 -- окончание миофиламенты на плазмолемме; Z - полоска Z. Электронная микрофотография.
Проводящая мышечная ткань также состоит из сердечных миоцитов, которые ио сравнению с клетками рабочей мускулатуры имеют больший диаметр, грушевидную или удлиненную форму, богаты анастомозами. Их светлые ядра с малым количеством гетерохроматина и хорошо выраженным ядрышком локализуются в центре клетки. Цитоплазма богата гликогеном н бедна митохондриями, что свидетельствует об интенсивно протекающем в ней гликолизе и низком уровне окислительных процессов. Незначительно развиты рибосомы, саркоплазматическая сеть, система поперечных трубочек, мало миофибрилл. Последние занимают периферическую часть клетки и не имеют определенной ориентации, в связи с чем Слабо выражена поперечная исчерченность. Так как миоциты содержат мало миоглобулина и внутриклеточных структур, то они окрашиваются слабее, чем клетки рабочей мускулатуры (рис. 149).
Между собой кардиомиоциты проводящей
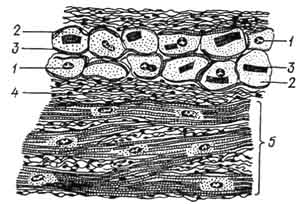
Рис. 149. Клетки проводящей мышечной ткани сердца быка:
A - продольное, Б - поперечное сечение; 1 - ядро; 2 - цитоплазма; 3 - миофибриллы; 4 - саркоплазма; 5 - рабочая мускулатура.
мускулатуры соединяются при помощи десмосом, а также щелевидных коп-тактов, которые создают возможность прямого контакта ионов.
Эта разновидность сердечной мышечной ткани формирует систему, обеспечивающую проведение возбуждения.
Отзывов (0)<