Гипофиз - компонент единой гипоталамофизарной системы организма. Вырабатывает гормоны, регулирующие функцию многих желез внутренней секреции и осуществляет их связь с центральной нервной системой. Расположен он в гипофизарной ямке турецкого седла клиновидной кости черепа; имеет бобовидную форму и очень небольшую массу. Так, у крупного рогатого скота она около 4 г, а у свиней меньше - 0,4 г.
Развивается гипофиз из двух эмбриональных зачатков, растущих навстречу друг другу. Первый зачаток - гипофизарный карман - образуется из крыши первичной ротовой полости и направлен в сторону головного мозга. Это эпителиальный зачаток, из которого в дальнейшем развивается аденогипофиз.
Второй зачаток - выпячивание дна мозгового желудочка, поэтому он является мозговым карманом и из него образуется нейрогипофиз (рис. 217).
Эмбриогенез обусловил строение органа - гипофиз состоит из двух долей: аденогипофиза и нейрогипофиза (рис. 218, 219).
В состав аденогипофиза входят передняя, промежуточная и туберальная части. Передняя часть построена из эпителиальных клеток - аденоцитов, формирующих тяжи (трабекулы) и разграниченных синусоидными капиллярами вторичной сосудистой сети (цв. табл. VII - А - а). Первичная сосудистая сеть расположена в медиальной эминенции.
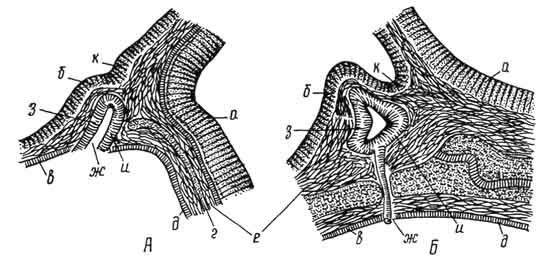
Рис. 217. Развитие гипофиза:
А - ранняя и Б - более поздняя стадии; а - стенки нервной трубки; б - стенки мозгового пузыря; в - эпителий ротовой бухты; г - хорда; д - кишечная трубка; е - мезенхима; ж - гипофизарный карман; его з - передняя и и - задняя стенки; к - зачаток задней части.
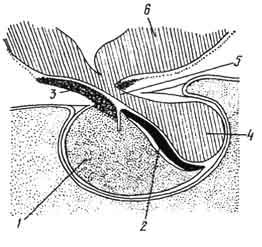
Рис. 218. Строение гипофиза:
1 - передняя, 2 - промежуточная, 3 - туберальная и 4 - задняя части; 5 -воронка; 6 - гипоталамус.
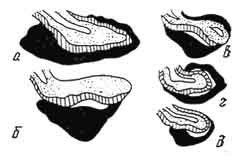
Рис. 219. Схема медиального разреза гипофиза домашних животных:
а - лошади; б - крупного рогатого скота; в - свиньи; г - собаки; д - кошки (по Траутману и Фибигеру).
Соединительнотканная строма аденогипофиза развита слабо.
Аденоциты по-разному воспринимают красители: клетки, хорошо окрашивающиеся, называют хромофильными, а плохо красящиеся - хромофобными (б). Хромофильные аденоциты могут воспринимать или кислые красители или основные, поэтому первые именуют ацидофильными (б), вторые - базофильными (г).
Ацидофильные клетки составляют 30 - 35% всех клеток передней части гипофиза. Они имеют округлую или овальную форму, крупнее хромофобных и мельче базофильных аденоцитов. В цитоплазме ацидофила содержатся гранулы, окрашивающиеся эозином; ядро расположено в центре клетки. К нему прилегают комплекс Гольджи, незначительное количество крупных митохондрий, хорошо развита зернистая эндоплазматическая сеть, что свидетельствует об интенсивном синтезе белка.
В связи с разной гормонообразовательной функцией и строением, цитоплазматической зернистостью различают три разновидности ацидофильных аденоцитов: соматотропоциты, лактотропоциты, кортикотропоциты. Соматотропоциты продуцируют соматотропный гормон, стимулирующий рост тканей и всего организма в целом. Лактотропоциты образуют пролактин (лактотропный гормон), регулирующий процесс лактации и функциональное состояние желтого тела яичника. Кортикотропоциты вырабатывают кортикотропин, повышающий гормонообразовательную функцию коры надпочечников.
Секреторные гранулы соматотропоцитов шаровидной формы, в диаметре от 200 до 400 нм (рис. 220). Лактотропоциты имеют более крупные секреторные гранулы овальной формы с длиной 500 - 600 нм, шириной 100 - 120 нм. Секреторные гранулы кортикотропоцитов снаружи покрыты мембраной пузырчатой формы с плотной сердцевиной.
Базофильные аденоциты составляют 4 - 10% всех клеток передней части гипофиза. Это самые крупные клетки аденогипофиза. Их секреторные гранулы имеют гликопротеидный характер, поэтому окрашиваются основными красителями. Различают две разновидности этих клеток: гонадотропные и тиротропные. Гонадотропные клетки продуцируют фолликулостимулирующий гормон, регулирующий развитие женских и мужских половых клеток, секрецию половых органов самки и лютеинизирующего гормона, стимулирующего рост и развитие желтого тела в яичниках и
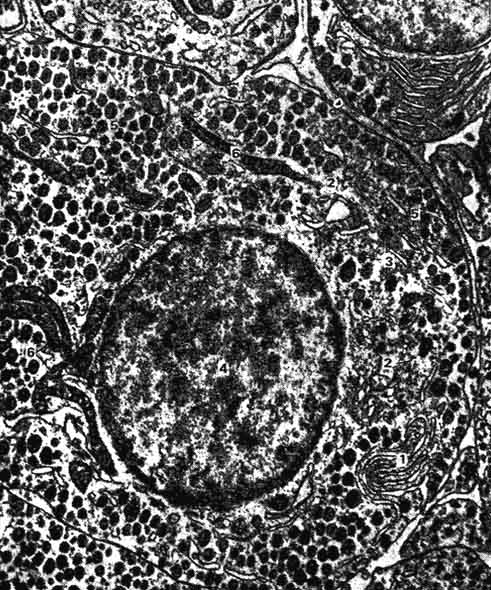
Рис. 220. Соматотропоцит передней доли аденогипофиза (электронная микрофотография):
1 - гранулярная эндоплазматическая сеть; 2 - комплекс Гольджи; 3 - Формирующиеся гранулы секрета; 4 - ядро; 5 - зрелые гранулы секрета; 6 - ? ;митохондрия (по Стрижкову).
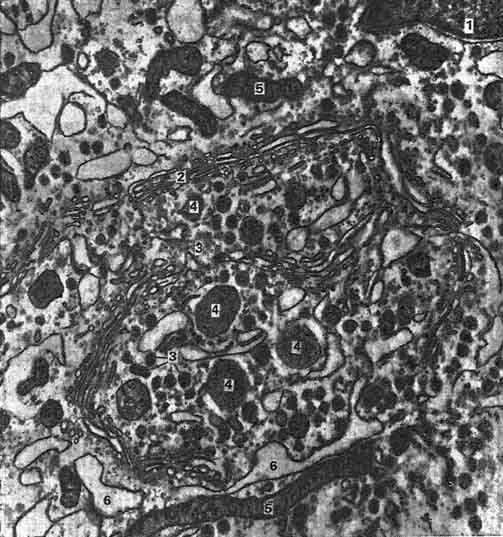
Рис. 221. Гонадотропоцит передней доли аденогипофиза:
1 - ядро; 2 - комплекс Гольджи; 3 - секреторные гранулы; 4 - накопительные гранулы; 5 - митохондрии; 6 - цистерны гранулярной эндоплазматической сети.
интерстициальных клеток в семенниках (рис. 221). В центральной зоне гонадотропного базофила находится макула. Это расширенная полость комплекса Гольджи, оттесняющая ядро, многочисленные мелкие митохондрии, мембраны эндоплазматической сети на: периферию клетки. Базофильные гонадотропоциты содержат зернистость, равную около 200 - 300 нм в диаметре.
При недостаточности половых гормонов в организме диаметр зернистости увеличивается. После кастрации животных базофильные гонадотропоциты превращаются в клетки кастрации: крупная вакуоль занимает всю центральную часть клетки. Последняя приобретает кольцевидную форму.
Тиреотропные базофилы (рис. 222) - угловатые клетки с мелкой (80 - 150 нм) заполняющей всю цитоплазму зернистостью. Если
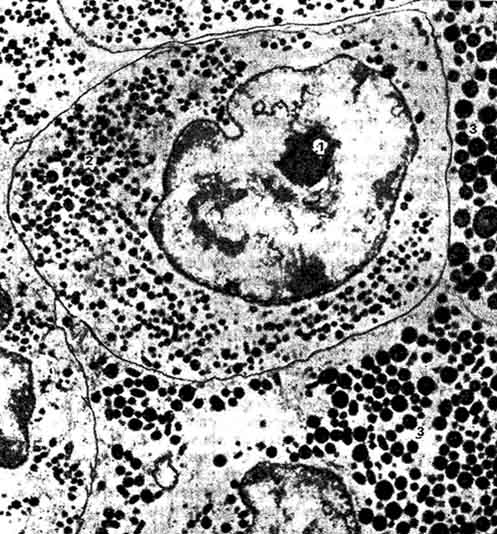
Рис. 222. Тиреотропоцит передней доли аденогипофиза (электронная микрофотография):
1 - ядро; 2 - секреторные гранулы; 3 - соматотропоцит (по Долан и Сёлоши).
организм испытывает недостаток гормонов щитовидной железы, то развиваются клетки тиреодэктомии. Они увеличены в размере, с расширенными цистернами эндоплазматической сети, поэтому цитоплазма имеет ячеистый вид, более крупные гранулы секрета,
Хромофобные клетки составляют 60 - 70% всех клеток передней части гипофиза. Это сборная группа, так как в ее состав входят разные по значению клетки: камбиальные, клетки на разной стадии дифференциации; не накопившие еще специфическую зернистость; клетки, выделившие секрет. Из камбиальных клеток в дальнейшем развиваются ацидофильные и базофильные аденоциты.
Промежуточная часть аденогипофиза представлена несколькими рядами слабобазофильных клеток. Продуцируемый
аденоцитами секрет накапливается в промежутках между клетками, что способствует образованию фолликулоподобных структур. Клетки промежуточной части аденогипофиза полигональной формы, содержат мелкие гликопротеидные гранулы размером 200 - 300 ни. В промежуточной зоне синтезируются меланотропин, регулирующий пигментный обмен, и липотропин - стимулятор жирового обмена.
Туберальная часть аденогипофиза по своей структуре сходна с промежуточной частью. Она прилегает к гипофизарной ножке и медиальной эминенции. Клетки этой зоны характеризуются слабой базофилией и трабекулярным расположением. Функция туберальной части окончательно не выяснена.
Выше говорилось о том, что гормонообразовательная функция аденогипофиза регулируется гипоталамусом, с которым он образует единую гипоталамоаденогипофизарную систему. Морфофункционально эта связь проявляется в следующем: верхняя гипофизарная артерия в медиальной эминенции образует первичную капиллярную сеть. Аксоны мелких нейросекреторных клеток ядер медиобазального гипоталамуса на сосудах первичной капиллярной сети образуют аксоваскулярные синапсы. Нейрогормоны, продуцируемые этими нейросекреторными клетками, по их аксонам перемещаются в медиальную эминенцию. Здесь они накапливаются, а затем через аксоваскулярные синапсы поступают в капилляры первичной сосудистой сети. Последние собираются в портальные вены, которые направляются вдоль гипофизарной ножки в аденогипофиз. Здесь вновь они распадаются и образуют вторичную капиллярную сеть. Синусоидные капилляры этой сети оплетают трабекулы секретирующих аденоцитов.
Кровь, оттекающая по венам от вторичной сосудистой сети, содержит аденогипофизарные гормоны, которые через общий кровоток, то есть гуморальным способом, регулируют функции эндокринных желез периферического звена.
Нейрогипофиз (задняя доля) развивается из мозгового кармана, поэтому он построен из нейроглии. Его клетки - питуициты веретенообразной или отростчатой формы. Отростки питуицитов контактируют с кровеносными сосудами. В заднюю долю входят крупные пучки нервных волокон, образованные аксонами нейросекреторных клеток паравентрикулярных и супраоптических ядер передней зоны гипоталамуса. Нейросекрет, образовавшийся этими клетками, перемещается вдоль аксонов в нейрогипофиз в виде секреторных капель. Здесь они оседают в виде накопительных телец, или терминалей, которые контактируют с капиллярами.
Следовательно, гормоны нейрогипофиза - окситоцин и вазопрессин синтезируются не структурами нейрогипофиза, а в паравентрикулярных и супраоптических ядрах. Затем, как говорилось выше, по нервным волокнам гормоны поступают в нейрогипофиз, где они накапливаются и откуда поступают в кровяное русло. Поэтому нейрогипофиз и гипоталамус тесно связаны и формируют единую гипоталамонейрогипофизарную систему.
Окситоцин стимулирует функцию гладких мышц матки, способствует выделению секрета маточных желез; при родах вызывает сильное сокращение мышечной оболочки стенки матки; регулирует сокращение мышечных элементов молочной железы.
Вазопрессин суживает просвет кровеносных сосудов и повышает кровяное давление; регулирует водный обмен, так как влияет на обратное всасывание (реабсорбцию) воды в канальцах почки.