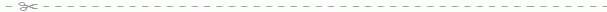Дробление - дальнейший процесс развития одноклеточной зиготы, в ходе которого образуется многоклеточная бластула, которая состоит из стенки - бластодермы и полости - бластоцеля. В бластодерме различают крышу, дно и расположенную между ними краевую зону. В процессе митотического деления зиготы образуются новые клетки - бластомеры, остающиеся тесно связанными друг с другом.
В начальной стадии дробления многоклеточный организм по своему размеру сходен с зиготой, так как его бластомеры, делясь, не достигают размера исходной клетки. Характер дробления в. эволюционном ряду хордовых различен, что в значительной степени обусловлено количеством и распределением желтка в яйцеклетках.
Дробление может быть полным (голобластическим) или частичным (меробластическим). При голобластическом дроблении принимает участие весь материал зиготы, при меробластическом - только та ее зона, которая лишена желтка.
Полное дробление классифицируют на равномерное и неравномерное. Полное равномерное дробление (рис. 43) характерно для яиц с малым количеством желтка (олиголецитальных) и равномерно расположенным по всей цитоплазме клетки желтком (изолецитальных). Примером такого дробления могут служить ланцетник, аскарида и др. В оплодотворенной яйцеклетке различают два полюса: верхний - анимальный и нижний - вегетативный.
После оплодотворения желток, незначительное количество которого было равномерно распределено то всей цитоплазме, перемещается к вегетативному полюсу. Первая борозда дробления проходит в меридиональном направлении и делит зиготу на два бластомера, которые соответствуют будущей левой и правой половине тела зародыша. Вторая борозда дробления проходит также меридионально под прямым углом к первой, и теперь зародыш состоит из четырех бластомеров. Третья борозда дробления имеет экваториальное направление, поэтому каждый бластомер делится на две части. Такой зародыш построен из восьми бластомеров, при этом четыре из них образовались из вегетативного полюса зиготы, в связи с чем они содержат весь желток зиготы и отличаются большими размерами. Эти бластомеры соответствуют задней части тела; анимальные - четыре - передней части.
Затем появляются две меридиональные борозды, делящие зародыш на 16 бластомеров. Пятое дробление - это две широтные борозды, в составе зародыша 32 бластомера. Они начинают
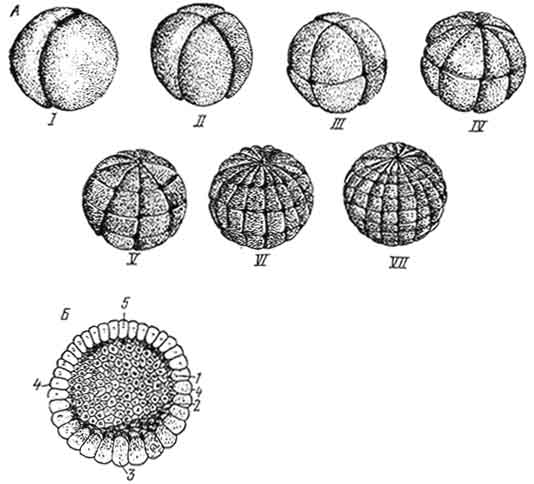
Рис. 43. Схема расположения борозд дробления у ланцетника (A):
I - зародыш на стадии двух бластомеров; II - зародыш на стадии четырех бластомеров; III - зародыш на стадии восьми бластомеров; IV - зародыш на стадии 16 бластомеров; V - зародыш на стадии 32 бластомеров; VI - зародыш на стадии 64 бластомеров; VII - зародыш на стадии 128 бластомеров. Строение бластулы (Б): 1 - бластодерма; 2 - бластоцель; 3 - дно; 4 - краевая зона; 5 - крыша бластулы.
постепенно отодвигаться друг от друга, контактируя лишь боковыми поверхностями. Внутри зародыша образуется сначала небольшая полость - бластоцель, которая постепенно увеличивается. После шестого дробления образуется 64 клетки, при этом борозды дробления проходят меридионально. После седьмого дробления (возникают четыре широтные борозды) зародыш состоит из 128 бластомеров.
Позднее синхронность в делении зародыша нарушается, бластомеры отодвигаются на периферию и располагаются в один слой, формируя бластодерму, а в центре зародыша образуется бластоцель.
Дробление завершается образованием бластулы, форма которой напоминает шар, заполненный жидкостью. Стенка шара образована клетками бластодермы.
Таким образом, при полном равномерном дроблении материал всей зиготы участвует в делении и после каждого деления (дробления) число клеток (бластомеров) увеличивается вдвое.
В бластодерме дифференцируются следующие участки: крыша, построенная из относительно мелких бластомеров; дно - это более крупные бластомеры и краевая зона, лежащая между дном и крышей бластулы.
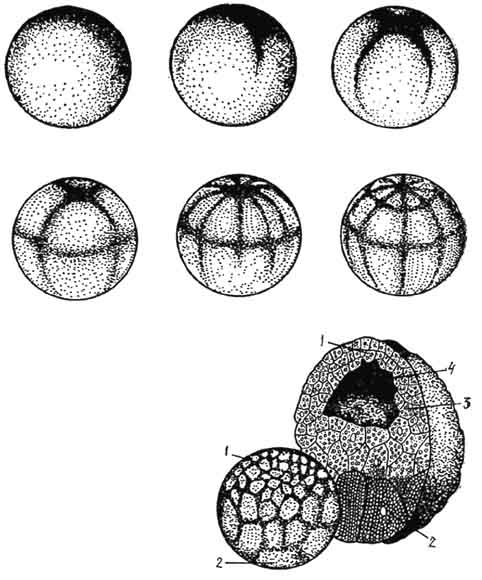
Рис. 44. Полное неравномерное дробление зиготы амфибии. Строение бластулы:
1 - микромеры; 2 - макроциты; 3 - бластодерма; 4 - бластоцель.
Полное неравномерное дробление характерно для мезолецитальных (среднее количество желтка) и телолецитальных (желток расположен в вегетативном полюсе) яйцеклеток. Примером этого типа дробления может служить дробление зиготы амфибий (рис. 44).
Дробление начинается с образования двух меридиональных борозд дробления, следующих друг за другом под прямым углом. Они быстро делят лишенный желтка анимальный полюс зиготы на два, а затем на четыре мелких бластомера. Вегетативный полюс, содержавший весь желток зиготы, дробится значительно медленнее, и бластомеры, возникающие здесь, более крупных размеров.
Третья борозда проходит ближе к анимальному полюсу зиготы и имеет широтное направление. Широтные борозды дробления сменяются меридиональными, при этом очень скоро возникает асинхронность и тангенциальность (деление бластомеров в
Рис. 45. Частичное (дискоидальное) дробление зародыша курицы:
А, В - стадии дробления - вид сверху (А - две меридиональные борозды, В - более поздняя стадия дробления); С - разрез зародышевого диска (а, b, с, - краевые клетки, расположенные на желтке; d, e, f, g, h - клетки, изолированные от желтка).
плоскости, параллельной поверхности зиготы) в дроблении, поэтому оно завершается образованием многослойной бластулы. Крыша бластулы построена из мелких бластомеров, именуемых микромерами. Дно состоит из крупных бластомеров - макромеров. Весь желток локализован в макромерах. Бластоцель сдвинут к анимальному полюсу и уменьшен в размере. Бластула, образовавшаяся в процессе голобластического (полного) дробления, носит название целобластулы.
Частичное, или меробластическое (дискоидальнее), дробление распространено у рыб, рептилий, птиц и характерно для полилецитальных (много желтка) и телолецитальных яиц (рис. 45).
В дроблении участвует только лишенный желтка поверхностный слой анимального полюса зиготы, так как здесь находятся ядро клетки и цитоплазма без желтка. Вся остальная часть зиготы загружена желтком и поэтому не дробится.
Первые две меридиональные борозды проходят через анимальный полюс под углом один к другому. Они не распространяются на вегетативный полюс, в связи с чем последний остается неразделенным на бластомеры. Меридиональные борозды сменяются широтными и тангенциальными. Бластомеры, образовавшиеся к ходе дробления, располагаются на желтке в один слой. Этот слой называется зародышевым диском, поэтому дробление получило название дискоидального.
На построение тела зародыша используется только его центральная часть - зародышевый щиток. Остальная часть зародышевого диска участвует в образовании временных (провизорные) органов - зародышевых оболочек, которые создают благоприятные условия для развития зародыша.
Дробление завершается образованием бластулы, у которое бластоцель имеет вид узкой щели и сдвинут к анимальному полюсу. Крыша бластулы построена из бластомеров. Краевая зона - это интенсивно делящиеся клетки (бластомеры) периферической зоны зародышевого диска. Дном является неразделенный на бластомеры желток вегетативного полюса зиготы. Такой тип бластулы называется дискобластулой.
Таким образом, из приведенного материала следует, что у хордовых имеется определенная зависимость между количеством желтка в яйцеклетках и характером дробления. Оно изменяется от полного (голобластического) к частичному (меробластическому), а бластула - от целобластулы к диско бластуле.
Общими свойствами развивающихся зародышей всех классов животных на стадии дробления являются постепенное увеличение числа клеток, а следовательно, и ДНК, так как дочерние клетки всегда диплоидные; увеличение площади клеточных поверхностей; возрастание региональных различий клеточных популяций.