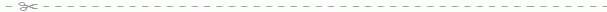Концевые аппараты нервных волокон - нервные окончания - различны по их функциональному значению. Различают три вида нервных окончаний: эффекторные, рецепторные и концевые аппараты в составе межнейрональных синапсов.
Эффекторные нервные окончания - к их числу относят двигательные нервные окончания поперечнополосатых и гладких мышц и секреторные окончания железистых органов.
Двигательные нервные окончания поперечнополосатых скелетных мышц - моторные бляшки - комплекс взаимосвязанных структур нервной и мышечной тканей. Моторная бляшка - эффекторный аппарат аксонов нервных клеток двигательных ядер передних рогов спинного мозга или двигательных ядер головного мозга и мышечных волокон. Морфологически он состоит из нервного полюса - терминальной части аксона нейрона и мышечного полюса, специализированного участка мышечного волокна - подошвы моторной бляшки (рис. 166).
Моторное нервное волокно вблизи мышечного волокна утрачивает сопровождающие осевой цилиндр ядра глиальных клеток и миелиновую оболочку. Осевой цилиндр, распадаясь на несколько терминальных веточек, погружается в специализированную шипу мышечного волокна.
Сакролемма в области нервного окончания образует многочисленные субмикроскопические складки, формирующие вторичные синаптические щели моторного окончания.
Мышечное волокно в области подошвы моторной бляшки не имеет миофибрилл и поперечной
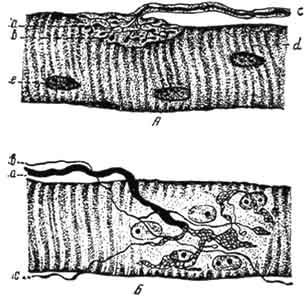
Рис. 166. Двигательное нервное окончание (моторная бляшка):
А - вид в профиль (а и b - окончания миелинового нервного волокна, с - миелиновое волокно, d - мышечное волокно, е - ядро мышечного волокна); В - вид сверху (а - миелиновое волокно, b - безмиелиновое нервное волокно, с - волокно, выходящее из моторной бляшки и оканчивающееся в другой моторной бляшке, так называемое "ультратерминальное волокно").
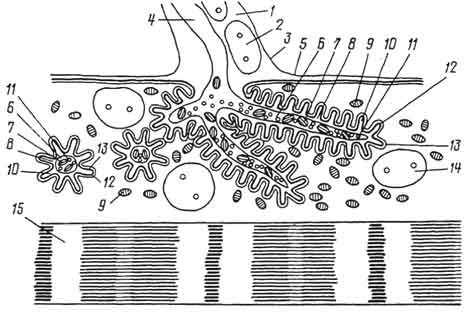
Рис. 167. Схема строения моторной бляшки:
1 - цитоплазма леммоцита; 2 - ядро; 3 - неврилемма; 4 - осевой цилиндр; 5 - сарколемма; 6 - концевые веточки нервного волокна в продольном и поперечном сечениях; 7 - митохондрии в нейроплазме (аксоплазме); 8 - первичное синаптическое пространство; 9 - саркосомы; 10 - вторичное синаптическое пространство; 11 - синаптические пузырьки; 12 - пресинаптическая мембрана; 13 - постсинаптическая мембрана; 14 - ядро моторной бляшки (мышечное); 15 - миофибрилла, состоящая из миопротофибрилл.
исчерченности. Здесь цитоплазма содержит значительное количество митохондрий и круглых или овальных ядер. Совокупность этих структур мышечного волокна в области нервного окончания образует его мышечный полюс.
Терминальные ветви осевого цилиндра нервного волокна характеризуются наличием митохондрий и многочисленных синаптических пузырьков, содержащих медиатор - ацетилхолин (рис. 167). Последний при деполяризации плазмолеммы аксона - пресинаптической мембраны - поступает в синаптическую щель и на холинорецепторы постсинаптической мембраны, которой служит оболочка мышечного волокна, что вызывает возбуждение (волну деполяризации постсинаптической мембраны).
Моторные нервные окончания гладкой мышечной ткани формируются нервными волокнами, которые распространяются между мышечными клетками и образуют четкообразные расширения, содержащие холинергические или адренергические пузырьки.
Чувствительные нервные окончания (рецепторы) - специализированные концевые образования дендритов чувствительных нейронов. В соответствии с их локализацией и специфичностью участия в нервной регуляции жизнедеятельности организма различают две большие группы рецепторов: экстерорецепторы и пптерорецепторы. В зависимости от характера воспринимаемого раздражения чувствительные окончания делятся на механорецепторы, хеморецепторы, терморецепторы и др.
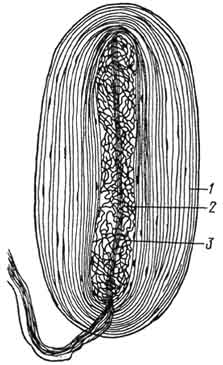
Рис. 168. Пластинчатое тельце (тельце Фатера - Пачини):
1 - наружная колба; 2 - внутренняя колба; 3 - концевой отдел нервного волокна (по Клара).
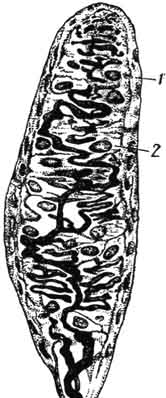
Рис. 169. Осязательное (мейсиерово) тельце:
1 - капсула; 2 - специальные клетки.
Чувствительные нервные окончания весьма разнообразны по их структурной организации. Они подразделяются на свободные нервные окончания, состоящие только из терминальных ветвей дендрита чувствительной клетки, и несвободные, содержащие в своем составе клетки глии. Несвободные окончания, покрытые соединительнотканной капсулой, называются инкапсулированными. Примером свободных нервных окончаний могут служить терминальные ветвления дендритов чувствительных клеток в эпидермисе кожи, где чувствительные нервные волокна, проникая в эпителиальную ткань, распадаются на тонкие терминальные веточки.
Весьма разнообразны чувствительные окончания в соединительной ткани животных, которые представлены двумя группами: неинкапсулированных и инкапсулированных нервных аппаратов. Первые содержат ветвящийся осевой цилиндр волокна, сопровождающий глии. Вторые характеризуются наличием соединительнотканной капсулы и специфичностью-= морфологии и функций их глиальных элементов. К группе таких чувствительных окончаний относят пластинчатые тельца (тельца Фатера - Пачини), осязательные тельца (тельца Мейснера), генитальные тельца и др. (рис. 168, 169).
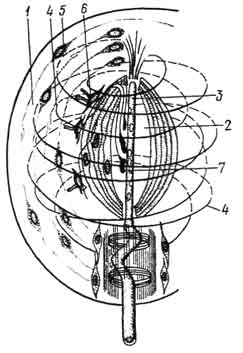
Рис. 170. Схема строения пластинчатого тельца:
1 - слоистая капсула; 2 - внутренняя колба; 3 - дендрит чувствительной нервной клетки; 4 - спиральные коллагеновые волокна; 5 - фиброциты; 6 - глиальные клетки с ресничками; 7 - синаптические контакты аксонов вторично чувствующих клеток с дендритами чувствительной нервной клетки (по Отелину).
В составе пластинчатого тельца различают внутреннюю колбу и капсулу. Внутренняя колба образована специализированными леммоцитами. В нее погружен осевой цилиндр - терминальный отдел чувствительного нервного волокна. Проникая во внутреннюю колбу, он распадается на тончайшие конечные ветвления.
Капсула пластинчатого тельца состоит из большого числа соединительнотканных пластинок, образованных фибробластами и спирально ориентированными пучками коллагеновых волокон. На границе наружной капсулы и внутренней колбы лежат клетки, которые предположительно определяют как глиальные. Они образуют синапсы с ветвлениями осевого цилиндра (рис. 170). Предполагают, что нервный импульс генерируется в условиях смещения наружной капсулы по отношению к внутренней колбе. Пластинчатые тельца характерны для глубоких слоев кожи и внутренних органов.
Осязательные тельца также образованы клетками глии, которые ориентированы перпендикулярно к длинной оси тельца, и стелются по их поверхности терминальными ветвями аксона. С поверхности тельце покрыто тонкой соединительнотканной капсулой.
Аналогично построены и генитальные тельца половых органов. Отличительной особенностью этого вида окончаний является то, что в генитальное тельце под капсулу проникает не один осевой цилиндр, а несколько. Последние ветвятся между глиальными клетками тельца. По той же схеме построены колбы Краузе, с функцией которых связывают температурную чувствительность. При их возбуждении медиатор поступает в синаптическую щель на холинорецепторы постсинаптической мембраны мышечного волокна и вызывает импульс (волну деполяризации) .
Рецепторы скелетных мышц - мышечные веретена содержат несколько интрафузальных мышечных волокон, покрытых общей соединительнотканной капсулой. В состав веретена обычно входят два толстых центральных мышечных волокна и
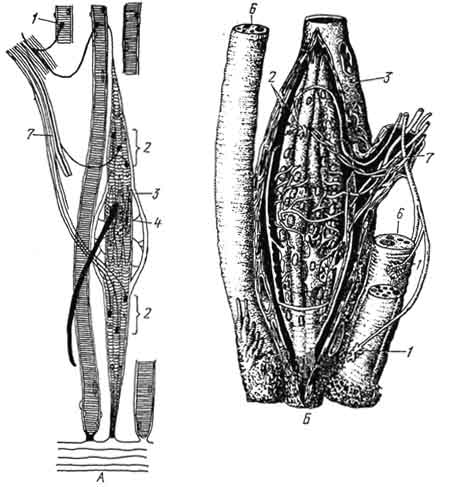
Рис. 171. Схема строения нервно-мышечного веретена:
А - моторная иннервация интрафузальных и экстрафузальных мышечных волокон (по Студитскому); Б - спиральные афферентные нервные окончания вокруг ннтрафузальных мышечных волокон в области ядерных сумок (по Кристичу с изменением); 1 - моторные бляшки экстрафузальных мышечных волокон; 2 - моторные бляшки интрафузальных мышечных волокон; 3 - ядерная сумка; 4 - ядерная сумка; 5 - чувствительные аннулоспиральные нервные окончания вокруг ядерных сумок; 6 - поперечнополосатые мышечные волокна; 7 - нерв.
четыре тонких (рис. 171). Экваториальная часть толстых волокон заполнена скоплениями ядер - "ядерная сумка". В тонких мышечных волокнах ядра располагаются цепочкой, образуется ядерная цепь. Чувствительные нервные волокна представлены здесь двумя видами. Одни образуют спиральные завитки, окружающие экваториальную, содержащую ядра, часть толстых интрафузальных мышечных волокон - "аннулярные окончания". Окончания второй группы чувствительных волокон представлены как аннулярными окончаниями, так и вторичными гроздевидными окончаниями по одному с каждой стороны от первичного. Окончания первой группы реагируют на степень растяжения мышц и на его скорость, вторичные - только на степень растяжения. На обоих полюсах мышечных волокон локализуются окончаниядвигательных нервных волокон и имеют строение, типичное для моторной бляшки.
Межнейрональный синапс - специализированный контакт двух нейронов, обеспечивающий одностороннее проведение нервного возбуждения. Морфологически в синапсе различают пресинаптический полюс - концевой отдел первого нейрона, и постсинаптический полюс - область контакта второго нейрона с пресинаптическим полюсом первого. Встречаются синапсы с химической и электрической передачей.
По месту контакта преганглионарного нервного волокна с вторым нейроном бывают аксосоматические синапсы (аксон первого нейрона контактирует с перикарионом второго), аксодендритические (аксон первого нейрона взаимодействует с дендритом второго) и аксоаксональные (аксон одного нейрона оканчивается на аксоне другого) (рис. 172). Предполагается, что последний не возбуждает нервного импульса на втором нейроне, а тормозит возбуждение, полученное нейроном через другие синапсы.
Морфологически пресинаптический полюс синапса характеризуется наличием синаптических пузырьков, содержащих медиатор (ацетилхолин или норадреналин), митохондрий, одиночных цистерн, иногда нейротубул. Участие в передаче нервного импульса следующему нейрону определяется закономерным выбросом путем экзоцитоза в синаптическую щель медиатора.
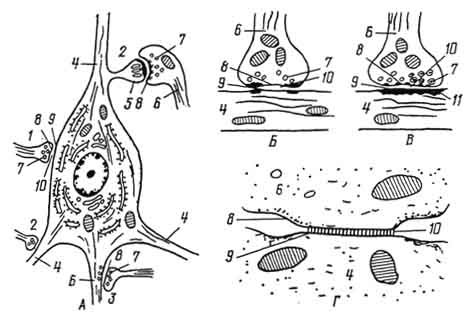
Рис. 172. Схема ультрамикроскопического строения различных типов синапсов:
А - цитотопография синапсов; Б - синапс тормозного типа; В - синапс возбудительного типа; Г - электротонический синапс; 1 - аксосоматический синапс; 2 - аксодендритические синапсы; 3 - аксоаксональный синапс; 4 - дендриты; 5 - дендритный шипик; 6 - аксон; 7 - синаптические пузырьки; 8 - пресинаптическая мембрана; 9 - постсинаптическая мембрана; 10 - синаптическая щель; 11 - постсинаптические структуры.
Последний, поступая на мембрану постсинаптического полюса, вызывает изменение ее проницаемости, волну деполяризации - генерацию нервного импульса. Роль медиатора, кроме указанных выше, могут играть и другие вещества, а именно: адреналин, серотонин, гамма-аминомасляная кислота и др.
Синаптические пузырьки, несущие различные медиаторы, морфологически различны. В холинэргических синапсах они: мелкие (30 - 40 нм) и прозрачные. В их составе иногда содержится также несколько очень крупных и электроноплотных пузырьков (80 - 150 нм), химический состав и значение последних в настоящее время неясны. Предположительно считают, что они содержат биогенные амины. Синаптические пузырьки адренэргических синапсов крупнее (50 - 90 нм) и морфологически характеризуются наличием в них электроноплотной гранулы. Выделение медиатора происходит экзоцитозом в синаптическую щель содержимого синаптических пузырьков.
Постсинаптическая мембрана холинэргических синапсов содержит "холинрецептивный" белок. При взаимодействии с ацетилхолином происходят конформационные изменения его молекул, приводящие к изменению проницаемости мембран и генерации нервного импульса в нейроне (рис. 173). Медиатор тормозных синапсов - гамма-аминомасляная кислота не увеличивает проницаемость постсинаптической мембраны для ионов, а уменьшает ее и, следовательно, стабилизирует мембранный потенциал, то есть тормозит генерацию нервного импульса.
На синаптических мембранах имеются характерные специализации. Под мембранами как пресинаптического, так и постсинаптического полюсов отмечают скопления плотного материала и тонких филаментов. Скопления обычно толще на
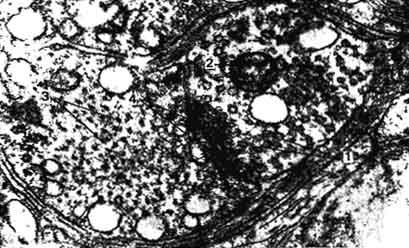
Рис. 173. Электронная микрофотография аксодендригического синапса в краниальном шейном узле кота (препарат Козлова):
1 - Синаптические пузырьки; 2 - митохондрия; 3 - нейротрубочки в цитоплазме дендрита; 4 - утолщенная постсинаптическая мембрана.
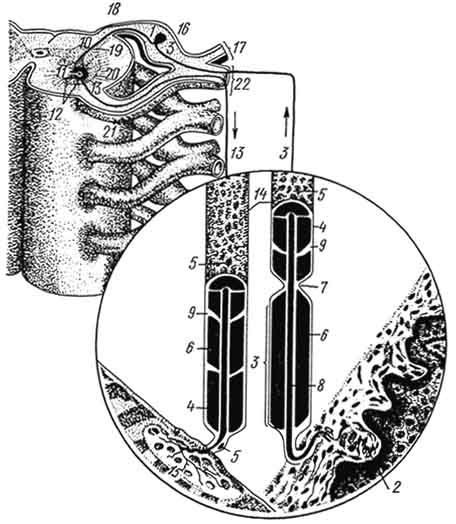
Рис. 174. Простая рефлекторная дуга:
1 - чувствительная нервная клетка; 2 - рецептор в коже; 3 - дендрит чувствительной клетки; 4 - оболочка; 6 - ядро леммоцита; 6 - миелиновая оболочка; 7 - перехват нервного волокна; 8 - осевой цилиндр; 9 - насечка на нервном волокне; 10 - нейрит чувствительной клетки; 11 - двигательная клетка; 12 - дендрит двигательной клетки; 13 - нейрит двигательной клетки; 14 - миелиновые волокна; 15 - эффектор (моторная бляшка); 16 - спинномозговой узел; 17 - дорсальная ветвь спинномозгового нерва; 18 - задний корешок; 19 - задний рог; 20 - передний рог; 21 - передний корешок; 22 - вентральная ветвь спинномозгового нерва.
пресинаптической мембране. С пресинаптическими уплотнениями часто связаны синаптические пузырьки. На тангенциальных срезах уплотнении видно, что они не гомогенны, а состоят из шестиугольных и треугольных структур, в центре которых можно различить синаптический пузырек. Скопления пузырьков и уплотнения вместе называют синаптическим комплексом, а поскольку они, по-видимому, являются местами преимущественного скопления пузырьков и выделения медиатора, их называют также активными зонами. В области синапсов выявляют и маленькие прикрепительные устройства - точки прикрепления (punctum adherens). От уплотнений синаптических комплексов они отличаются большей толщиной и симметричностью и малой линейной протяженностью.
Электротонические синапсы образуются при плотном прилежании плазмолемм, двух нейронов, преимущественно их дендритов, и перикариона.
Нервная система организма представлена чувствительными, ассоциативными и двигательными клетками, объединенными межнейрональными синапсами в функционально активные образования - рефлекторные дуги. Простая рефлекторная дуга состоит из двух нейронов - чувствительного и двигательного (рис. 174).
В подавляющем большинстве рефлекторные дуги высших позвоночных животных содержат в своем составе еще значительное количество ассоциативных нейронов, расположенных между чувствительным и двигательным нейронами.