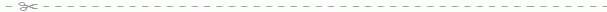Лейкоциты - разнообразные по морфологическим признакам и функциям клетки сосудистой крови. В организме животных они выполняют многообразные функции, направленные прежде всего на защиту организма от чужеродного влияния путем фагоцитарной активности, участия в формировании гуморального и клеточного иммунитета, а также в восстановительных процессах при тканевом повреждении.
В крови животных лейкоцитов в 600 - 800 раз меньше, чем эритроцитов. В 1 mkл крови у крупного рогатого скота их насчитывают 4,5 - 12,0 тыс., у лошадей - 7,0 - 12,0 тыс., овец - 6,0 - 14,0 тыс., свиней - 8,0 - 16,0 тыс., собак - 8,5 - 10,5 тыс., кур - 20,0 - 40,0 тыс. Увеличение количества лейкоцитов - лейкоцитоз - характерный признак для многих патологических процессов, но может наблюдаться и у здоровых животных (лейкоцптоз новорожденных, пищеварительный лейкоцитоз и т. д.).
Образовавшись в кроветворных органах и поступив в кровь, лейкоциты лишь непродолжительное время пребывают в сосудистом русле, затем они мигрируют в вокругсосудистую соединительную ткань и органы, где осуществляют свою основную функцию. Для многих лейкоцитов тканевая фаза их жизни является завершающей.
Особенность у лейкоцитов та, что они обладают подвижностью. В цитоплазме лейкоцитов, непосредственно около плазмолеммы, имеется комплекс актиновых микрофиламентов, с помощью которых приводится в движение клеточная поверхность и образуются псевдоподии. Изменяя свою внешнюю форму и форму ядра, белые кровяные клетки способны активно перемещаться между клетками эндотелия сосудов, проникать через базальные мембраны и клетки эпителия, мигрировать в основном веществе соединительной ткани.
В лейкоцитах различают ядро и цитоплазму, содержащую различные органеллы и включения. Классификация лейкоцитов основана на учете их морфологических признаков, выявляемых при световой микроскопии окрашенных мазков крови и имеет прежде всего клинико-практическое значение. Те лейкоциты, в цитоплазме которых содержится специфическая зернистость, называются зернистыми (или гранулоцитами). Зрелые зернистые лейкоциты, как правило, имеют расчлененное на сегменты ядро - сегментоядерные клетки. В соответствии с различием в окрашивании цитоплазматической зернистости в группе гранулоцитов выделяют три вида клеток: нейтрофилы - зернистость окрашивается и кислыми, и основными красителями; эозинофилы - зернистость окрашивается кислыми красителями и базофилы - зернистость окрашивается основными красителями. Незернистые лейкоциты (агранулоциты) характеризуются отсутствием специфической зернистости в цитоплазме и несегментированными ядрами. В группе агранулоцитов выделяют два вида - лимфоциты и моноциты. Таким образом, схема классификации лейкоцитов выглядит следующим образом:

В ветеринарной практике при анализе крови животных важное диагностическое значение имеет дифференциальный подсчет лейкоцитов. Определенное процентное соотношение между отдельными видами лейкоцитов называют лейкоцитарной формулой - лейкограммой. При определении лейкоцитов на окрашенных мазках крови с помощью светового микроскопа учитывают многие морфологические признаки: величину клеток, форму и плотность ядра, соотношение между объемом ядра и цитоплазмы, наличие или отсутствие в цитоплазме зернистости, ее окраску, величину и распределение в объеме цитоплазмы (рис. 96). В настоящее время лейкоцитарную формулу устанавливают с помощью автоматических приборов.
Гранулоциты (зернистые лейкоциты).
Нейтрофнльные лейкоциты (нейтрофилы) - очень подвижные клетки, обладающие высокой фагоцитарной активностью.
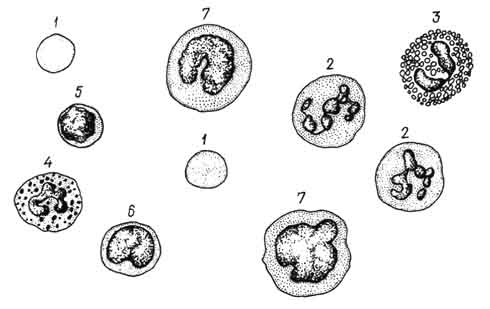
Рис. 96. Клетки крови животных при световой микроскопии:
1 - эритроциты; 2 - нейтрофилы; 3 - эозинофил; 4 - базофил; 5 - малый лимфоцит; 6 - средний лимфоцит; 7 - моноциты.
2. Лейкограмма крови животных. %
И. И. Мечников назвал их микрофагами. Нейтрофилы всего лишь несколько часов циркулируют в сосудистой крови. Благодаря хемотаксису - направленному движению фагоцитов под влиянием хемотаксических факторов нейтрофилы мигрируют из кровеносных капилляров в соединительную ткань, накапливаются в очаге воспаления, где и осуществляют свою основную фагоцитарную функцию, обеспечивая очищение очага воспаления от микроорганизмов и продуктов клеточного и тканевого распада.. В процессе фагоцитоза нейтрофилы погибают и вместе с бактериальными веществами и остатками разрушенных тканей образуют массу, называемую гноем. Хемотаксическими свойствами обладают многие продукты экзо- и эндогенного происхождения и прежде всего вещества, выделяемые бактериями и вирусами, самими-лейкоцитами, а также образующиеся при тканевом распаде.
В крови животных нейтрофилов содержится от 25 до 70% всех лейкоцитов. Больше всего нейтрофилов у лошадей, северных оленей, собак - до 65 - 70%. Это животные с так называемым нейтрофильным профилем крови. Нейтрофилез - увеличение в лейкоцитарной формуле процента нейтрофилов - наиболее типичен для гнойно-воспалительных процессов.
В мазках крови, окрашенных но Романовскому - Гимзе, размер нейтрофилов от 8 до 15 мкм, их определяют главным образом по форме и структуре ядра. У зрелых нейтрофилов ядро состоит из нескольких сегментов, соединенных тонкими перемычками.. Очень большая сегментированность ядер характерна для нейтрофилов крови овец. У этих животных часто встречаются клетки с 8 - 10 сегментами. В сегментах содержится плотный примембранный хроматин, благодаря чему они интенсивно окрашиваются. В крови животных (см. лейкограмму) имеется определенное количество менее зрелых предшественников сегментированных форм - палочкоядерных нейтрофилов, имеющих ядро в виде изогнутой палочки или буквы S и метамиелоцитов (юных), содержащих бобовидное ядро. Соотношение этих трех видов нейтрофилов имеет определенное диагностическое значение в ветеринарной клинике. При многих патологических состояниях уменьшается количество сегментоядерных и нарастает количество палочкоядерных и юных нейтрофилов, мобилизуемых из красного костного мозга в сосудистую кровь. Обогащение крови более молодыми формами нейтрофилов получило название "сдвига влево", так как при выведении лейкограммы незрелые клетки принято записывать слева, а зрелые - справа. Резкий сдвиг влево наблюдают при тяжелых септических инфекциях.
Цитоплазма нейтрофилов слабооксифильна, в ней содержится очень мелкая, трудно различимая при световой микроскопии -окрашенных мазков крови зернистость. Отношение зернистости к красителям варьирует у разных животных: у собаки, кошки и свиньи зернистость окрашивается в розово-фиолетовый цвет. У кроликов гранулы окрашиваются эозином в ярко-красный цвет, то есть имеют выраженное сродство к кислым красителям. По этой причине нейтрофилы кроликов иногда называют псевдоэозинофилами. Оксифильна и палочковидная зернистость нейтрофилов у птиц.
При электронно-микроскопическом исследовании в цитоплазме нейтрофилов обнаруживают небольшое количество органелл общего назначения, включения гликогена, в периферической зоне содержатся микрофиламенты, обеспечивающие движение клетки. Характерным для цитоплазмы нейтрофилов является наличие в ней гранул двух разновидностей: азурофильных и специфических, содержимое которых позволяет этим клеткам выполнять свои функции. Появляющиеся на ранних стадиях развития более крупные и электроноплотные азурофильные гранулы содержат, кроме типичных гидролитических ферментов, миелопероксидазу и лизоцим. В многочисленных (до 80% всего количества гранул), но более мелких специфических гранулах выявляется щелочная фосфатаза. Показано, что с фагосомой первой по времени контактирует и сливается специфическая гранула, в последующем переваривание осуществляется с помощью ферментов азурофильных гранул. Дегрануляция нейтрофилов и активизация лизосомальных гидролитических ферментов происходит и при разрушении клеток, что наблюдается при воспалительной реакции. Таким образом, благодаря совместному действию многочисленных ферментов, содержащихся в гранулах, нейтрофилы могут переваривать все макромолекулы, присутствующие в бактериях и очаге воспаления.
Эозинофильные лейкоциты (эозинофилы) - разновидность гранулоцитов. Количество их у животных составляет 2 - 12% всех лейкоцитов сосудистой крови. Наибольший процент эозинофилов в крови крупного рогатого скота, верблюдов и овец. По размеру эозинофилы крупнее нейтрофилов и имеют диаметр 12 - 18 мкм. Ядро в зрелых эозинофилах также сегментированное, но чаще ядро состоит из двух сегментов. Значительную сегментированность ядра (3 - 5 сегментов) отмечают в эозинофилах овец. В крови животных встречаются палочковидные и юные формы эозинофилов.
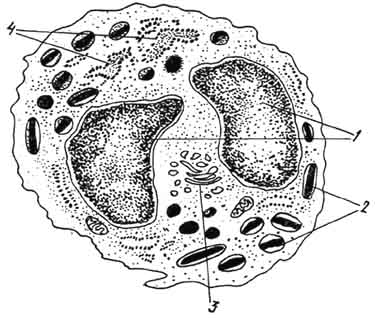
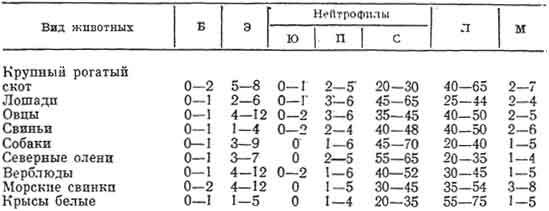
Рис. 97. Схема ультрамикроскопического строения эозинофильного гранулоцита крысы:
1 - сегменты ядра; 2 - зрелые специфические гранулы, содержащие кристаллоиды; 3 - комплекс Гольджи; 4 - гранулярная эндоплазматическая сеть (рис. Козлова).
Весьма характерна для эозинофилов относительно крупная специфическая зернистость цитоплазмы. Гранулы, как правило, равномерно и плотно заполняют площадь цитоплазмы. Зерна ярко окрашиваются в оранжево-красный или красный цвет, то есть имеют выраженную оксифилию вследствие содержания в них основных белков. Особенно крупные размеры гранул в эозинофилах однокопытных (лошадь, осел). Гистохимическими методами в зернистости выявлены различные ферменты: кислая фосфатаза, пероксидаза, гистаминаза, арилсульфатаза и др.
При электронной микроскопии зрелые гранулы эозинофилов некоторых видов животных имеют расположенные в центре электроноплотные палочковидные структуры, окруженные по периферии более светлым содержимым (рис. 97).
Подобно нейтрофилам, эозинофилы обладают хемотаксисом. Хемотаксическими веществами в отношении эозинофилов являются комплексы антиген - антитело, гистамин и другие низкомолекулярные факторы, выделяемые тканевыми базофилами. По сравнению с нейтрофилами эозинофилы менее подвижны и обладают меньшей фагоцитарной активностью. Во время фагоцитоза происходит дегрануляция эозинофилов, высвобождение из гранул многих ферментов, с помощью которых осуществляется инактивация гистамина и других медиаторов воспаления. Таким образом, важная функция эозинофилов - антигистаминовая, участие в ограничении воспалительного процесса.
Эозинофилы являются важнейшими эффекторными клетками в противопаразитарном иммунитете. Многие паразитарные болезни животных (фасциолез, эхинококков и др.) сопровождаются усилением продукции эозинофилов в красном костном мозге и увеличением их количества в сосудистой крови (у крупного рогатого скота до 20%). В антипаразитарном механизме важнейшее значение имеет прямой контакт эозинофилов с возбудителем, поэтому они в больших количествах скапливаются вокруг внедрившихся в организм паразитов. Характерно содержание значительного количества эозинофилов в рыхлой соединительной ткани стенка кишечника.
Базофильные лейкоциты - базофилы - самая малочисленная разновидность гранулоцитов периферической крови животных (0,5 - 2% всех лейкоцитов). В крови сельскохозяйственных птиц 3 - 4% базофилов, а содержание их в крови земноводных доходит до 20%. В мазках окрашенной крови диаметр базофилов несколько меньший, чем у других гранулоцитов, - 10 - 12 мкм. Ядро чаще в виде неправильно изогнутой палочки, реже сегментированное. Слабооксифильная цитоплазма содержит неравномерно распределенную, различную по величине и форме, легко растворимую в воде зернистость. При окраске по Романовскому - Гимзе зернистость красно-вишневого цвета, то есть обладает метахромазией - окрашивается в цвет, отличающийся от цвета основного красителя (азура). Это свойство метахромазии обусловлено особой формой полимеризации молекул красителя при его взаимодействии с гепарином, входящим в состав гранул. Кроме гепарина, в базофилах имеются гранулы, содержащие высокоактивное вещество - гистамин. Электронно-микроскопически гранулы неодинаковы по электронной плотности (рис. 98).
На поверхности базофилов расположены специальные рецепторы, с помощью которых связываются иммуноглобулины E. Присоединение антигенов (аллергенов) и образование комплекса антиген - антитело вызывают дегрануляцию базофилов и выход значительного количества содержащихся в них фармакологически активных веществ - гистамина и гепарина. Быстрое высвобождение этих соединений и лежит в основе реакций повышенной чувствительности немедленного типа - сывороточной болезни, феномена Артюса и др. Установлено, что базофилы составляют значительную часть клеточного инфильтрата при различных формах гиперчувствительности замедленного типа. Таким образом, базофилы участвуют в иммунологических реакциях организма, в частности в реакциях аллергического типа.
Агранулоциты (незернистые лейкоциты). Моноциты. В 1 мкл крови у животных содержится от 1 до 8%. Моноциты периферической крови - самые крупные из лейкоцитов - в мазках окрашенной крови их диаметр 18 - 20 мкм и более. Это клетки с относительно большим количеством голубовато-серой цитоплазмы (с мелкой азурофильной зернистостью) и различным по своей форме ядром. Последнее чаще всего бобовидной или лопастной формы, в нем различают расположенный преимущественно по периферии конденсированный хроматин и мелкие ядрышки. При
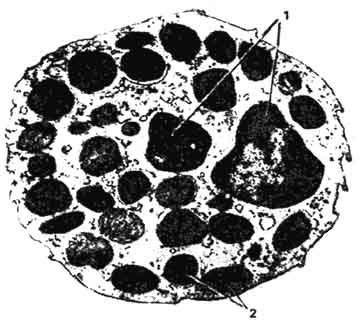
Рис. 98. Электронная микрограмма базофильного гранулоцита:
1 - сегменты ядра; 2 - базофильные гранулы (по Блюму и Фаусету).
электронной микроскопии в цитоплазме выявляются лизосомы, пиноцитозные пузырьки, развитый комплекс Гольджи, небольшие по размеру митохондрии и плотные гранулы, соответствующие азурофильным гранулам, видимым в световой микроскоп (рис.99). Цитохимическими методами в моноцитах обнаруживают липиды, гликоген; в них высокая активность неспецифической эстеразы, имеются кислые гидролазы, пероксидаза, лизоцим (мурамидаза), с участием которых обеспечивается антимикробное действие. По цитохимической характеристике моноциты имеют сходство с клетками нейтрофильного ряда.
Циркулирующие в крови моноциты являются предшественниками тканевых и органных макрофагов, на основании чего они отнесены к системе мононуклеарных фагоцитов (СМФ). После пребывания в сосудистой крови (12 - 36 ч) моноциты мигрируют через эндотелий капилляров и венул в ткани и превращаются в фиксированные и подвижные макрофаги. При этом в них отмечается сильное развитие лизосом и появление фагосом. В большом количестве моноциты выходят из кровяного русла в ткани при воспалительных реакциях, в том числе и иммунных. В очагах хронического воспаления моноциты становятся важнейшим клеточным элементом, выполняющим многообразные функции (стимулируют фибробластические процессы, образуют многоядерные клетки и др.).
Лимфоциты - важнейшие клетки, участвующие в разнообразных иммунологических реакциях организма. Это многочисленная группа лейкоцитов крови животных. У крупного рогатого скота,
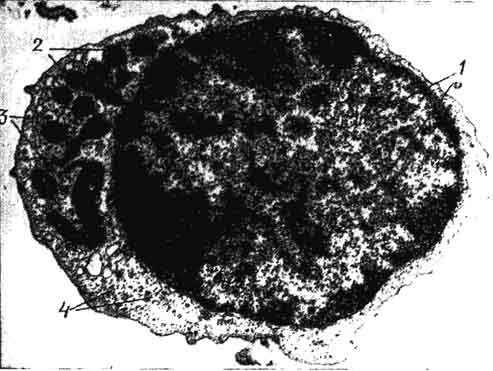
Рис. 99. Электронная микрограмма среднего лимфоцита:
1 - ядро; 2 - митохондрии; 3 - мелкие канальцы гранулярной эндоплазматической сети; 4 - свободные полисомы.
овец, кроликов, белых крыс число лимфоцитов составляет 40 - 65% всех лейкоцитов. У других животных (лошади, собаки, свиньи, северные олени), отличающихся преобладанием в крови нейтрофилов, лимфоцитов содержится 20 - 40%. Большое количество лимфоцитов находится в лимфе.
По размеру и некоторым структурным особенностям, выявляемым при световой микроскопии окрашенных мазков крови, различают малые (диаметр до 8 мкм), средние (8 - 11 мкм) и большие (более 11 мкм) лимфоциты. Малые лимфоциты составляют основную массу (до 90%) всех лимфоцитов крови животных. Это клетки с выраженным преобладанием объема ядра над объемом цитоплазмы. Ядро у них чаще всего круглое с небольшим углублением. Крупные конгломераты конденсированного хроматина расположены как по периферии, так и в центральной части ядра, что делает его темным и маскирует ядрышко. Базофильная цитоплазма обнаруживается или в виде узкого вокругядерного ободка, или в виде серпа, прилежащего к одной стороне ядра. В цитоплазме некоторых лимфоцитов содержится небольшое количество мелких азурофильных гранул.
Для средних и больших лимфоцитов более характерны ядра слегка бобовидной формы с меньшим количеством конденсированного хроматина и отчетливо заметным ядрышком, а также болееширокий ободок цитоплазмы. Электронно-микроскопически в цитоплазме малого лимфоцита выявляют свободные полисомы и рибосомы. Другие органеллы - митохондрии, мелкие канальца эндоплазматической сети, элементы комплекса Гольджи, как правило, немногочисленны и располагаются преимущественно наряду с центриолями около ядерного углубления.
Морфологически сходные малые лимфоциты крови являются функционально разнородными клетками, развитие которых до их поступления в сосудистую кровь происходит в различных органах.
Различают два основных класса лимфоцитов: T- и В-лимфоциты.
Т-лимфоциты развиваются из костномозговых клеток в корковой части долек тимуса. Их дифференцировка выражается в появлении в плазмолемме поверхностных антигенных маркеров (антигены гистосовместимости) и многочисленных рецепторов, с помощью которых происходит распознавание чужеродных антигенов и иммунных комплексов.
В-лимфоциты свое название получили потому, что у птиц этот вид лимфоцитов образуется из стволовых предшественников в фабрициевой сумке (Bursa - В). Местом развития В-лимфоцитов у животных считают миелоидную ткань костного мозга. В ходе дифференцировки В-лимфоциты приобретают способность синтезировать иммуноглобулины, которые становятся составными элементами их плазмолеммы и выполняют роль рецепторов. С их участием В-лимфоциты взаимодействуют с антигенами при их распознавании. Кроме того, на поверхности В-лимфоцитов образуются рецепторы для связывания комплемента (его компонента С3) и рецепторы для Fc фрагмента иммуноглобулиновых молекул.
Установлено, что в основном (более 70%) малые лимфоциты сосудистой крови относятся к T-лимфоцитам, среди которых значительное количество долгоживущих клеток памяти (10 лет и более). Большинство В-лимфоцитов относится к короткоживущим (недели - месяцы).
Циркулирующие в крови T- и В-лимфоциты представляют подвижную популяцию относительно функционально неактивных клеток. Вне сосудистого русла они скапливаются в определенных участках периферических лимфоидных органов (для Т-лимфоцитов это тимусзависимые зоны), где могут при действии соответствующих антигенов подвергаться активации, превращаться в бластные формы, размножаться и дифференцироваться в эффекторные клетки клеточного и гуморального иммунитета, а также создавать увеличенный фонд клеток памяти.
Эффекторными клетками в системе Т-лимфоцитов являются три основные субпопуляции: Т-киллеры (цитотоксические лимфоциты), Т-хелперы (помощники) и Т-супрессоры (угнетающие). Эффекторными клетками В-лимфоцитов являются плазмобласты и зрелые плазмоциты, способные в повышенном количестве продуцировать иммуноглобулины.
Субпопуляции Т-системы также отличаются своими поверхностными маркерами, биологическими свойствами и выполняют различные специфические иммунологические функции, которые составляют основу двух различных иммунных ответов - клеточного и гуморального.
Цитотоксические лимфоциты (Т-киллеры - убийцы) путем непосредственного контактного воздействия или с помощью близкодействующих токсических медиаторов (лимфокинов) разрушают чужеродные клетки-мишени или изменившиеся собственные клетки. Такой тип реакции и обезвреживания антигенов (не сопровождающийся выработкой растворимых антител) называют клеточным иммунитетом. Он наблюдается при отторжении трансплантатов, при реакциях гиперчувствительности замедленного типа, при разрушении опухолевых клеток и развитии аутоиммунных реакций.
Т-хелперы и Т-супрессоры - две субпопуляции клеток, имеющих значительную роль в регуляции активности В-лимфоцитов. Т-хелперы выделяют продукты своей синтетической деятельности (медиаторы), которые, взаимодействуя с В-лимфоцитами, способствуют их превращению в антителопродуцирующие плазматические клетки. Т-супрессоры подавляют процессы деления В-клеток и образования плазмоцитов и таким образом оказывают тормозящий эффект на антителообразование. Согласно современным представлениям, в механизме действия клеток хелперов и супрессоров важнейшее значение отводится Fc рецепторам, находящимся на поверхности В-клеток. Вспомогательный эффект Т-хелперов рассматривают как блокировку этих Fc рецепторов медиаторами хелперов, что препятствует связыванию рецепторов иммуноглобулинами, но способствует их связыванию со специфическими антигенами. Медиаторы Т-супрессоров способствуют образованию связи между иммуноглобулинами и Fc рецепторами В-клеток, тем самым ингибируют их превращение в плазмоциты. В тех случаях, когда антиген вызывает в организме образование антител, В-лимфоциты находятся под влиянием Т-хелперов и макрофагов. Показано, что для развития гуморального иммунного ответа на тимусзависимые антигены превращение В-лимфоцита в плазмоцит не может происходить без участия всех трех типов клеток: Т-хелперов, В-лимфоцитов и макрофагов. Имеется ряд гипотез о том, каким образом происходит взаимодействие клеток в процессе этого иммунного ответа.
На часть тимуснезависимых антигенов (пневмококковый полисахарид, липополисахарид кишечной палочки и др.) В-лимфоциты отвечают самостоятельно. Этим антигенам свойственна особая структура молекул, позволяющая им концентрироваться на поверхности В-клеток без помощи Т-хелперов.
Существенная функция T-лимфоцитов - их способность синтезировать и выделять ряд факторов белковой природы - лимфокинов, которые изменяют поведение других клеток (фактор, угнетающий миграцию макрофагов, фактор активации макрофагов, хемотаксические факторы для нейтрофилов, эозинофилов, базофилов), обладают противовирусной активностью (интерферон, лимфотоксин) и т. д.
Основными методами разграничения и выявления морфологически сходных, но функционально разнородных лимфоцитоподобных T- и В-клеток и их субпопуляций в настоящее время являются иммунологические, основанные на определении рецепторов клеточной поверхности. Для идентификации В-лимфоцитов по поверхностным иммуноглобулинам используется метод иммунофлуоресценции. Идентификация Т-лимфоцитов in vitro основана на их способности к спонтанному розеткообразованию с эритроцитами барана (метод бласттрансформации с фитогемаагглютинином или конканавалином А).